Гаструляция у ланцетника представляет удобную модель для изучения общих закономерностей этого процесса у типа хордовых. Этот процесс, как было показано ранее, у ланцетника, происходит по типу инвагинации. На ранней стадии гаструлы обнаруживается два зародышевых листка: эктодермальный, образованный из крыши бластулы, и энтодермальный- из дна бластулы. Гастроцель сообщается с внешней средой при помощи бластопора.
Наступает следующий этап развития зародыша - образование третьего зародышевого листка-мезодермы. Клетки дорзальной губы бластопора, быстро размножаясь, погружаются в составе энтодермы и на дорзальной ее стороне образуют хордальную пластинку- зачаток будущей хорды. Мелкие клетки вентральной и боковых губ бластопора также врастают в энтодермальный листок и располагаются в его составе по сторонам от хордальной пластинки.
Двуслойный зародыш приобретает вытянутую форму, имея в своем составе материал трех зародышевых листков. Материал мезодермы занимает дорзальную часть внутреннего листка. Из первичной эктодермы, расположенной над хордальной пластинкой, выделяется нервная пластинка. Последняя погружается и по длине зародыша образует нервный желобок. Нервный желобок, замыкаясь кверху, образует нервную трубку.
Хордальная пластинка выходит из состава первичного внутреннего листка, замыкается книзу и образует плотный клеточный тяж - хорду.
Одновременно мелкоклеточный материал, находившийся в первичном внутреннем листке по сторонам от хорды, выпячивается в дорзальном направлении в виде двух карманов. Вначале эти карманы открываются в гастроцель, а затем отделяются в виде двух замкнутых складок, расположенных вдоль гаструлы.
Так образуется третий зародышевый листок - мезодерма. После выделения хорды и мезодермы материал первичного внутренного листка смыкается, образуя энтодерму - внутренний зародышевый листок (рис. 5).
ГАСТРУЛЯЦИЯ у птиц
Гаструляция у птиц представляет довольно близкую модель процессам, имеющим место у высших хордовых, в том числе и у человека.
У птиц в результате дискоидального (меробластического) дробления образуется бластодиск. Он состоит из клеток неправильной формы, плотно прилегающих друг к другу и расположенных на огромной массе нераздробившегося желтка. Под влиянием ферментов клеток бластодиска часть желтка разжижается и образуется полость, заполненная жидкостью - бластоцель. Крыша этой бластулы представлена бластодиском, дно - нераздробившейся массой желтка. Бластоцель - подзародышевая полость.
Затем наступает гаструляция, состоящая из двух фаз. Первая фаза начинается до откладывания яйца и заключается в обособлении энтодермы путем деляминации клеток бластодиска. В результате возникает двуслойный зародыш. Верхний слой клеток дискобластулы приобретает призматическую форму и располагается в один правильный ряд. Нижний слой клеток дискобластулы сохраняет округлую или неправильную форму и лежит беспорядочно на желтке. Между клетками верхнего и нижнего слоя возникает полость - гастроцель.
Вторая фаза гаструляции начинается во второй половине первых суток инкубации. Бластодиск разрастается, в центре его выделяется зародышевый щиток, из которого в дальнейшем развивается тело зародыша. Окружающая зародышевый щиток часть дискобластулы представляет внезародышевый материал, в котором различают светлое и темное поле. Светлое поле расположено вокруг зародышевого щитка и состоит из клеток, отделившихся от желтка подзародышевой полостью в силу частичного использования желтка зародышем. Темное поле занимает периферию дискобластулы и состоит из клеток, плотно прилегающих к желтку и растущих по его поверхности. К этому моменту резко выражено разделение клеточного материала на зародышевую (зародышевый щиток) и внезародышевую (светлое и темное поле) часть.
В течение 24 часов насиживания или инкубации на зародышевом щитке в результате размножения клеток наблюдается перемещение их спереди назад по периферии зародышевого щитка. Оба потока клеток встречаются в центре у заднего конца зародышевого щитка, сливаются и перемещаются кпереди посредине зародышевого щитка. В результате образуется утолщенный тяж клеток, получивший название первичной полоски.
На переднем конце первичной полоски формируется утолщение- первичный узелок. В дальнейшем часть бластомеров, расположенных кпереди от первичного узелка, перемещается к последнему, подворачивается под эктодерму и формирует растущий кпереди между эктодермой и энтодермой головной или хордальный вырост - зачаток хорды.
Бластомеры задней половины зародышевого щитка смещаются к первичной полоске и через эту область погружаются под эктодерму, располагаясь в полости между эктодермой и энтодермой. Этот материал образует зачаток мезодермы - третьего зародышевого листка. Клетки хордального выроста размножаются и. перемещаются, занимая центральное положение между эктодермой

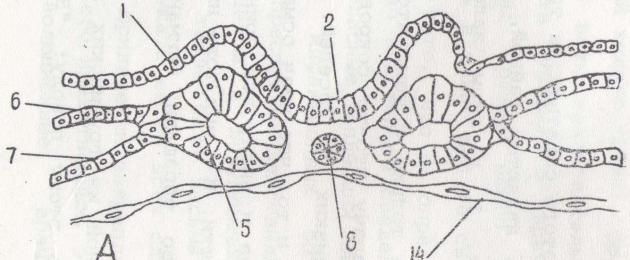
Рис.%6. Развитие комплекса осевых органов у птиц 1-эктодерма; 2 -нервный желобок; 3 - нервная трубка; 4 - амнион; 5 - сомиты; 6 - париетальный листок сплахнотома; 7 - висцеральный листок спланхнотома; 8 - хорда; 9 - дерматом; 10 - миотом; 11 - склеротом; 12 - нефротом; 13 - аорта; 14 - энтодерма; 15 - амниотическая складка и энтодермой в передней половине зародышевого щитка Вслед за этим клетки мезодермы также перемещаются в переднюю половину зародышевого щитка, располагаясь между эктодермой и энтодермой по бокам от хордального выроста. Это парахордальная мезодерма. Ее можно подразделить на дорзальную и вентральную.
По мере того как клетки первичной полоски уходят в закладки хорды и мезодермы на поверхности остается материал эктодермы и входящей в нее нервной пластинки. Таким образом, первичная полоска и головной узелок по своему значению соответствуют бластопору ланцетника, т. к. в этом месте проходит перемещение зародышевого материала, приводящее к образованию 3-зародышевых листков. Наиболее глубоко расположен зачаток кишечной энтодермы. Он непосредственно прилегает к поверхности желтка и находится во внутреннем слое дискобластулы.
К концу гаструляции в теле зародыша имеются все зачатки, из которых возникает типичный для хордовых комплекс осевых органов (рис. 6.).
Результат активного деления клеток, роста и направленных перемещений (миграций) клеточных потоков с формированием многослойного зародыша, или гаструлы, (возникновением послойно расположенных, отделенных друг от друга отчетливой щелью, зародышевых листков: наружного - эктодермы, среднего - мезодермы, внутреннего - энтодермы). Перемещение клеток происходит в строго определенной области зародыша - в области серого серпа. Последний был описан В. Ру в 1888 г. В оплодотворенном яйце амфибии серый серп выявляется как окрашенная область на стороне, противоположной проникновению спермия. В этом месте, как полагают, локализуются факторы, необходимые для гаструляции.
У разных представителей позвоночных совершается несколькими основными способами: путем инвагинации (впячивания), иммиграции (перемещения части клеток внутрь зародыша), эпиболии (обрастания), деламинации (расщепления). Способы гаструляции зависят от типа яйцеклетки. При любом способе гаструляции в качестве ведущих сил выступают неравномерная пролиферация клеток в разных частях зародыша, уровень обменных процессов в клетках, расположенных в разных частях зародыша, активность амебоидных движений клеток, а также индуктивные факторы (белки, нуклеопротеиды, стероиды и др.). В результате гаструляции обособляются основные зачатки органов и тканей.
Следующим периодом эмбриогенеза является гисто- и органогенез - дифференцировка различных тканей и органов организма из материала зародышевых листков и эмбриональных зачатков.
В результате гаструляции возникает многослойный зародыш . Несмотря на различные способы гаструляции после выделения материала зародышевых листков по оси зародыша находится материал хорды, который подстилает нервную пластинку, слева и справа от хорды располагается материал мезодермы. Все это характеризует осевой комплекс зачатков. В дальнейшем происходит формирование зачатков органов, представляющих собой пространственно локализованные группы стволовых клеток - источников развития тканей. Закономерности дифференцировки клеточного материала зачатков можно проследить в эмбриогенезе наиболее изученных животных.
Ланцетник. Развитие ланцетника.
Классическим объектом эмбриологических исследований ланцентник , подробно изученный А.О. Ковалевским. Ланцетник является представителем класса хордовых животных подтипа бесчерепных, размером до 8 см и обитает на песчаном дне в теплых морях. Свое название получил из-за формы, напоминающей ланцет (хирургический инструмент с обоюдоострым лезвием, современный скальпель).
Яйцеклетка ланцетника олиго- и изолецитальная, размером 110 мкм, ядро располагается ближе к анимальному полюсу. Оплодотворение наружное. Дробление зиготы полное, почти равномерное, синхронное и заканчивается образованием бластулы. В результате чередования меридианальных и широтных борозд дробления формируется однослойная бластула с полостью, заполненной жидкостью - бластоцелем. Бластула сохраняет полярность, ее дно представляет собой вегетативную, а крыша - анимальную части; между ними находится краевая зона.
При гаструляции происходит впячивание вегетативной части бластулы в анимальную. Впячивание постепенно углубляется и, наконец, формируется двустенная чаша с широко зияющим отверстием, ведущим в новообразованную полость зародыша. Такой способ гаструляции называется инвагинацией. Так бластула превращается в гаструлу. В ней материал зародыша оказывается дифференцированным на наружный листок - эктодерму, и внутренний - энтодерму. Полость чаши именуется гастроцелем, или полостью первичной кишки, которая сообщается с внешней средой посредством бластопора, что соответствует заднепроходному отверстию. В бластопоре различают дорсальную, вентральную и две латеральных губы. В результате инвагинации центр тяжести зародыша смещается, и зародыш поворачивается бластопором кверху. Постепенно происходит смыкание краев бластопора и удлинение зародыша. Топография клеток в составе губ бластопора определяет развитие разных частей зародыша. При гаструляции из внутреннего листка гаструлы обособляются хорда и мезодерма, которые располагаются между экто- и энтодермой. Завершается гаструляция образованием осевого комплекса зачатков и дальше - обособлением зачатков органов. Хорда индуцирует развитие нервной трубки из материала дорсальной эктодермы. Эта часть эктодермы утолщается, образуется нервная пластинка (нейроэктодерма), которая прогибается по средней линии и превращается в желобок.

Края желобка постепенно смыкаются в нервную трубку. Оставшаяся часть эктодермы - кожная, срастается над нервной трубкой. Однако на самом переднем и заднем концах зародыша нервная трубка некоторое время сообщается с внешней средой с помощью двух отверстий - нейропоров. В последующем происходит разделение мезодермы на спинные сегменты - сомиты, количество которых нарастает от 15 пар до 60-65 пар у взрослого ланцетника. Часть латерально расположенной мезодермы не сегментируется и расщепляется на наружный (париетальный) и внутренний (висцеральный) листки спланхнотома. Эти листки растут между экто- и энтодермой и, достигнув середины на вентральной стороне зародыша под кишечной трубкой, срастаются, формируя единую вторичную полость - целом. На переднем конце зародыша возникает углубление (ротовая бухта), растущее по направлению к переднему отделу кишечной трубки. При контакте эктодермы ротовой бухты и слепого конца кишечной трубки происходит апоптоз клеток и возникает сообщение кишки с внешней средой. Аналогичный процесс протекает на заднем конце зародыша. По бокам головного отдела зародыша также возникает контакт кожной эктодермы и кишечной энтодермы. В месте этого контакта происходит прорыв. Так полость передней кишки сообщается с внешней средой (формируется жаберный аппарат). После этого зародыш выходит из яйцевой оболочки во внешнюю среду в виде личинки.
Методы маркирования для изучения процессов миграции бластомеров позволили выделить определенные области зародыша на ранних стадиях развития (зиготы - бластулы), которые позднее развиваются в зародышевые листки и эмбриональные зачатки органов и тканей. Эти области были названы презумптивными (предполагаемыми) участками, или зачатками.
Развитие ланцетника впервые было изучено А. О. Ковалевским. Вопрос этот имеет большой интерес, так как анализ стадий развития примитивнейшего из современных хордовых животных дает некоторые основания для суждения о ранних этапах филогении хордовых. Кроме того, развитие ланцетника представляет упрощенную схематическую картину эмбрионального развития и других хордовых. На рис 4 и рис 5 изображены последовательные стадии эмбрионального развития ланцетника вплоть до формирования личинки. Дробление оплодотворенного яйца полное и почти равномерное: при образовании бластулы видно, что на ее нижней стороне, соответствующей вегетативной ("растительной") части яйца, клетки крупнее, чем на верхней. В силу этого внутренний слой следующей затем стадии гаструлы представлен более крупными клетками. Дробление происходит очень быстро. В эктодерме верхней стороны зародыша обособляется медуллярная пластинка , края которой свертываются, а затем и смыкаются. Возникающая таким путем нервная трубка сохраняет на переднем конце (через невропор) некоторое время сообщение с внешней средой, а на заднем конце (через нервно-кишечный канал) - с полостью гаструлы, т. е. с первичной кишкой . В дальнейшем нервнокишечный канал исчезает вовсе, а на месте невропора остается обонятельная ямка .
Из боковой пластинки развиваются брюшина, брыжейки (в которых в виде продольных каналов возникают основные кровеносные сосуды), мускулатура кишечника. Нефридиальные канальцы развиваются в виде пальцевидных выпячиваний стенок вторичной полости тела. Гонады развиваются как выпячивания той части стенок полости тела, которая соответствует месту разделения сомита и боковой пластинки гонотому. Рот образуется путем выпячивания первичной кишки на конце, противоположном гастропору (первичному рту), и встречного впячивания эктодермы. В месте встречи этих образований возникает прорыв. Закладка рта и жаберных щелей происходит асимметрично. Ротовое отверстие закладывается на нижней левой стороне зародыша. Левые жаберные щели (их 14) первоначально возникают на брюшной стороне, а затем перемещаются на правую сторону зародыша. Затем здесь появляется еще один ряд щелей (их 8), расположенных выше упомянутых ранее 14 щелей. Впоследствии нижний ряд щелей смещается на брюшную сторону и лишь после этого - на левую сторону тела. Число их при этом сокращается с 14 до 8. Число жаберных щелей с обеих сторон затем резко возрастает. Впоследствии рот смещается на брюшную сторону. Атриальная полость возникает первоначально в виде желобка на нижней поверхности тела. Формирующие этот желобок метаплевральные складки растут навстречу друг другу и, смыкаясь, образуют полость, открывающуюся наружу лишь в задней своей части, где упомянутые складки не срастаются. В целом личиночное развитие ланцетника длится около трех месяцев.
Позвоночные животные произошли от бесчерепных. Современным представителем подтипа бесчерепных является ланцетник (рис. 34). В развитии ланцетника мы видим наиболее простую схему зародышевого развития хордовых животных, которая в значительной степени усложнилась в процессе эволюции у позвоночных животных и особенно у человека.
Ланцетник - морское животное. Самка и самец выбрасывают половые клетки (яйца и сперматозоиды) непосредственно в воду, где и происходит оплодотворение и развитие зародыша.
Вслед за оплодотворением зигота вступает в период дробления (рис. 35); количество бластомеров быстро увеличивается (2, 4, 8, 16 и т. д.).

В процессе деления бластомеры постепенно отодвигаются от центра зародыша к периферии, образуя в центре все увеличивающуюся полость.
В связи с этим к концу периода дробления возникает типичная бластула, стенка которой образована одним слоем клеток (бластодерма), а полость ее (бластоцель) заполнена жидкостью. Следующий период (гаструляция) связан с инвагинацией, т. е. впячиванием одной (вегетативной) половины бластулы в другую (анимальную) * (рис. 35). В результате образуется гаструла ** , у которой имеется внутренний зародышевый листок (первичная энтодерма), гастроцель (полость первичной кишки) и бластопор (первичный рот).
* (В яйце ланцетника в одной из его половин содержится больше желтка, чем в другой. В результате изучения развития зародышей установлено, что часть яйца, снабженная большим количеством желтка, при дроблении образует ту половину бластодермы, которая впячивается в периоде гаструляции и образует внутренний зародышевый листок - энтодерму. Известно, что из энтодермы образуется пищеварительная и другие системы так называемой растительной (вегетативной) жизни. Поэтому и часть яйца, содержащая большее количество желтка, и образующаяся из него в результате дробления часть бластулы называются вегетативными частями. Из противоположной части яйца и соответствующей части бластулы развиваются эктодерма и далее органы анимальной (животной) части, в частности нервная система и др. Поэтому эти части яйца и бластулы именуются анимальными. )
** (Греч, gaster - желудок. Отсюда название "гаструла", чтобы подчеркнуть, что зародыш в этой стадии снабжен зачатком пищеварительной системы в виде первичной кишки. )
Бластоцель (первичная полость тела) в период инвагинации сохраняется некоторое время в виде узкой щели между наружным и внутренним зародышевыми листками, а затем исчезает. После сформирования гаструла начинает увеличиваться в длину; одновременно происходит концентрическое смыкание краев бластопора (первичного рта) * .
* (Бластопор (первичный рот), соединяющий первичную кишку с внешней средой, у одних животных на последующих этапах развития остается в качестве ротового отверстия (первичноротые животные), а у других становится заднепроходным (анальным) отверстием (вторичноротые животные). В последнем случае ротовое отверстие образуется на противоположном конце. К первичноротым относятся черви, моллюски и членистоногие, к вторичноротым - иглокожие и хордовые, в частности ланцетник и позвоночные животные, включая человека. )
Окончание гаструляции совпадает с началом периода обособления основных зачатков органов и тканей (рис. 36). В это время утолщенный дорсальный участок первичной эктодермы превращается в нервную пластинку, из которой возникает, проходя через стадию нервного желобка, нервная трубка (рис. 36, 37). Нервная трубка является зачатком нервной системы.


Другая часть наружного зародышевого листка при дальнейшем развитии замыкается над нервной трубкой и является зачатком кожного эпителия (эпидермиса).
Внутренний зародышевый листок претерпевает ряд изменений в клеточном составе, затем из него обособляются следующие образования: в области средней части его крыши - хордальная пластинка, из которой образуется зачаток хорды; в области латеральных частей крыши первичной кишки - карманообразные выпячивания, которые затем обособляются от стенки первичной кишки. Клеточный материал отшнуровавшихся карманообразных выпячиваний первичной кишки заполняет первичную полость тела (находящуюся между эктодермой и энтодермой) и представляет собой зачаток среднего зародышевого листка (мезодермы). В центре обособившегося участка оказывается отшнуровавшееся от полости первичной кишки пространство, являющееся вторичной полостью тела. Оставшаяся часть первичной энтодермы (дно первичной кишки) образует кишечную трубку, которая является зачатком вторичной (окончательной) кишки и из которой в дальнейшем возникает кишечный эпителий.
Таким образом, к концу гаструляции, в период обособления зачатков органов и тканей, нервная трубка оказывается расположенной на спинной стороне зародыша в срединном положении, а под ней последовательно расположены хорда и кишечная трубка. Окончательно выявляется билатеральная симметрия. Появляются мезодермальные карманы. Зачатки мезодермы с полостями внутри них врастают справа и слева в щель между кожной эктодермой и кишечной трубкой и соединяются под последней. В то же время мезодерма, вытянутая вдоль тела между кожной эктодермой, с одной стороны, хордой и кишечной трубкой, с другой стороны, разделяется на ряд отдельных участков (сегментов), расположенных по длине тела друг около друга. Сегментации подвергаются левый и правый мезодермальные карманы. Одновременно каждый мезодермальный мешок на всем протяжении делится на дорсальный участок (сомит) и вентральный участок (спланхнотом). Сомиты утрачивают полость, становятся плотными и служат исходным материалом для туловищной мускулатуры. Спланхнотомы сохраняют полость. Они остаются некоторое время отделенными друг от друга (в результате сегментации), а затем изолированные полости, заключенные в отдельных спланхнотомах, сливаются, так что образуется единая для всех сегментов тела целомическая полость (вторичная полость тела, целом). Материал стенок спланхнотомов является исходным материалом эпителиальной выстилки вторичной полости тела * .
* (Вторичная полость тела (целом) возникает в толще среднего зародышевого листка. Из нее в процессе эмбрионального развития образуются различные полости. У человека таковыми являются, в частности, брюшинная и плевральные полости и полость перикарда. Первичная полость тела (бластоцель) исчезает в процессе гаструляции и образования мезодермы. )
Развитие личинки ланцетника заканчивается появлением ротового и анального отверстий, жаберных щелей, формированием органов и др.
Особенности эмбрионального развития анамний изучаются на примере ланцетника, рыб и земноводных.
Яйцеклетки у ланцетника – первичные изолецитальные , оплодотворение совершается в воде, т.е. наружное. После оплодотворения образуется зигота, которая подвергается полному и равномерному дроблению – развитие голобластическое . Зигота делится вначале двумя последовательными митозами во взаимно перпендикулярных меридиональных плоскостях на четыре, затем экваториальной бороздой на восемь бластомеров и т.д. Плоскости дробления чередуются, и после седьмого деления возникает бластула типа целобластулы .
Бластомеры , образующие бластодерму различаются по своим размерам и качеству, т.к. происходит распределение разнокачественного материала цитоплазмы зиготы, которая подвергается внутренней дифференцировке. Возникшая целобластула состоит из крупноклеточных желточных бластомеров, образующих дно (будущая кишечная энтодерма), средних по размеру бластомеров, расположенных над ними дорсально – материал дорсального серпа (будущая хорда) и мелких бластомеров, окружающих дно бластулы, – материал центрального серпа (будущая мезодерма). Все это окружено эктодермой.
Используя метод прижизненной окраски, было установлено, что все перечисленные участки бластулы перемещаются путем подворачивания через губы бластопора, размещаются вокруг гастроцеля и создают основу для протекания органотипического периода развития ланцетника – периода дифференцировки тканей и органов.
Бластула имеет полость – бластоцель . Бластоцель заполняется жидкостью – продуктом жизнедеятельности клеток бластодермы.
Путем инвагинации , т.е. втягивания вегетативного полушария в анимальное, бластула преобразуется в гаструлу , стенка которой становится двухслойной и состоит из эктодермы снаружи и энтодермы внутри. Это первичные зародышевые листки.
В гаструле образуется полость первичного кишечника – гастроцель , которая сообщается с внешней средой при помощи бластопора . Вследствие перемещения центра тяжести в сторону анимального полюса зародыш поворачивается на 180° бластопором кверху и продолжает плавать в воде.
Позже зародыш удлиняется. Из первичной энтодермы в дорсальном направлении выделяется хордальная пластинка, а в дорсолатеральном две мезодермальные пластинки.Из первичной эктодермы по средней линии тела выделяется нервная пластинка, состоящая из более высоких клеток, чем остальная эктодерма. Нервная пластинка отшнуровывается от эктодермы и погружается под нее, превращаясь вначале в нервныйжелобок , а затем в нервнуютрубку , остальная кожная эктодерма смыкается над нервной трубкой. Одновременно с образованием нервной трубки хордальная пластинка преобразуется в круглый клеточный тяж – хорду , мезодермальные пластинки свертываются в полые трубки, лежащие между хордой и кожной эктодермой, а оставшаяся энтодерма смыкается во вторичную кишку . Так образуется комплекс осевых органов, характеризующих тип хордовых животных.
Мезодерма метамерно (от головы и хвостовой части зародыша) разделяется на сегменты, причем в хвостовой части зародыша сегментации не происходит. Кроме того, первые два сегмента развиваются самостоятельно, воспроизводя древнюю трехсегментную личиночную форму бесчерепных – диплеврулу . Каждый сегмент мезодермы, исключая два первых («древние») сегмента, разрастается в дорсовентральном направлении и подразделяется на три части: сомит (дорсально), спланхнотом (вентрально) и сегментную ножку между ними.
Сомиты дифференцируются на дерматом – кожный листок (латерально), склеротом – зачаток скелета (центрально) и миотом – мышечный листок (остаток после выделения первых двух). Из миотома впоследствии развивается скелетная (соматическая) мускулатура. Спланхнотом расщепляется на два листка: висцеральный (внутренний) и париетальный (пристеночный), между ними находится вторичная полость тела – целом . Из обоих листков спланхнотома выделяется ткань сетевидной формы – мезенхима , которая также образуется из склеротома и дермотома сомита. Мезенхима (эмбриональная соединительная ткань) заполняет все пространство между тремя зародышевыми листками. Из оставшейся части обоих листков спланхнотома возникает выстилка целома – мезотелий . Наконец, сегментная ножка преобразуется в нефрогонотом – эпителиальную выстилку выделительной системы и зачаток половой.
Периодом дифференцировки тканей и органов заканчивается личиночный период развития ланцетника, который длится примерно три месяца, и из личинки возникает половозрелое животное.