LECTURE No. 5.
VIROLOGY.
All viruses exist in two qualitatively different forms. Extracellular form – virion – includes all the constituent elements of the viral particle. Intracellular form - virus – can be represented by only one nucleic acid molecule, because Once inside the cell, the virion breaks down into its constituent elements. At the same time, the intracellular virus is a self-replicating form that is not capable of division. On this basis, the definition of a virus presupposes a fundamental difference between cellular forms of existence (bacteria, fungi, protozoa) that reproduce by division, and a replicating form that reproduces from viral nucleic acid. But the distinctive features of viruses from pro- and eukaryotes are not limited to this. The fundamental differences include:
1. the presence of one type of nucleic acid (DNA or RNA);
2. lack of cellular structure and protein synthesizing systems;
3. the possibility of integration into the cellular genome and synchronous replication with it.
The shape of the virion can be very different (rod-shaped, ellipsoidal, spherical, filamentous, sperm-shaped), which is one of the signs of the taxonomic affiliation of this virus.
The sizes of viruses are so small that they are comparable to the thickness of the cell membrane. The smallest (parvoviruses) are 18 nm in size, and the largest (variola virus) are about 400 nm.
The classification of viruses is based on the type of nucleic acid that forms the genome, which made it possible to distinguish two subkingdoms:
riboviruses– RNA-containing or RNA viruses;
deoxyriboviruses– DNA-containing or DNA viruses.
Subkingdoms are divided into Families, Subfamilies, Genera and Species.
When systematizing viruses, the following main criteria were identified: similarity of nucleic acids, size, presence or absence of supercapsid, type of symmetry of the nucleocapsid, characteristics of nucleic acids, polarity, number of strands in the molecule, presence of segments, presence of enzymes, intranuclear or cytoplasmic localization, antigenic structure and immunogenicity, tropism for tissues and cells, the ability to form inclusion bodies. An additional criterion is the symptomatology of the lesions, i.e. the ability to cause generalized or organ-specific infections.
According to the structural organization they distinguish simply organized (“naked”) And complexly organized (“dressed”) viruses.
The structure of a simple virion is arranged in such a way that viral nucleic acid, those. the genetic material of the virus is reliably protected by a symmetrical protein shell – capsid, the functional and morphological combination of which forms nucleocapsid.
The capsid has a strictly ordered structure, which is based on the principles of helical or cubic symmetry. It is formed by subunits that are identical in structure - capsomeres, organized in one or two layers. The number of capsomeres is strictly specific for each species and depends on the size and morphology of the virions. Capsomeres, in turn, are formed by protein molecules - protomers. They can be monomeric - composed of a single polypeptide or polymer - composed of several polypeptides. The symmetry of the capsid is explained by the fact that a large number of capsomeres are required for genome packaging, and their compact connection is possible only with a symmetrical arrangement of subunits. The formation of the capsid resembles the crystallization process and proceeds according to the principle of self-assembly. The main functions of the capsid are determined by the protection of the viral genome from external influences, ensuring the adsorption of the virion on the cell, the penetration of the genome into the cell as a result of the interaction of the capsid with cellular receptors, and determine the antigenic and immunogenic properties of virions.
The nucleocapsid follows the symmetry of the capsid. At spiral symmetry the interaction of nucleic acid and protein in the nucleocapsid occurs along one axis of rotation. Each virus with helical symmetry has a characteristic length, width and periodicity. Most viruses pathogenic to humans, including the influenza virus, have helical symmetry. The organization based on the principle of helical symmetry gives viruses a rod-shaped or filamentous shape. This arrangement of subunits forms a hollow channel, inside which a viral nucleic acid molecule is compactly packed. Its length can be many times greater than the length of the virion. The tobacco mosaic virus, for example, has a virion length of 300 nm, and its RNA reaches a value of 4000 nm. With this organization, the protein sheath better protects hereditary information, but requires more protein, because the coating consists of relatively large blocks. At cubic symmetry the nucleic acid is surrounded by capsomers, forming an icosahedron - a polyhedron with 12 vertices, 20 triangular faces and 30 angles. The organization of the virion according to this principle gives viruses a spherical shape. The principle of cubic symmetry is the most economical for the formation of a closed capsid, because To organize it, small protein blocks are used, forming a large internal space into which the nucleic acid fits freely.
Some bacteriophages have double symmetry, when the head is organized according to the cubic principle, and the process according to the principle of spiral symmetry.
It is typical for large viruses lack of constant symmetry.
An integral structural and functional component of the nucleocapsid are internal proteins, ensuring correct supercoiled genome packaging and performing structural and enzymatic functions.
The functional specificity of viral enzymes is determined by their location and the mechanism of formation. Based on this, viral enzymes are divided into virus-induced And virion. The former are encoded in the viral genome, the latter are part of the virions. Virion enzymes are also divided into two functional groups: enzymes of the first group ensure the penetration of viral nucleic acids into the cell and the exit of daughter populations; enzymes of the second group are involved in the processes of replication and transcription of the viral genome. Along with their own, viruses actively use cellular enzymes that are not virus-specific. But their activity can be modified during the process of virus reproduction.
There is a group of so-called. complexly organized or "dressed" viruses, which, unlike "naked", have a special lipoprotein shell on top of the capsid - supercapsid or peplos, organized by a bilayer of lipids and specific viral glycoproteins that penetrate the lipid bilayer and form spine-like growths(peplometers or supercapsid proteins ). Surface supercapsid proteins are an important component that facilitates the penetration of viruses into sensitive cells. It is these special proteins called F proteins ( fusio - fusion), ensures the fusion of viral supercapsids and cell membranes. The supercapsid is formed at the later stages of the reproductive cycle during the budding of daughter populations and is a derived structure from the membranes of a virus-infected cell. Thus, the composition of lipids depends on the nature of the “budding” of the viral particle. For example, the influenza virus has a lipid bilayer composition similar to that of cell membranes. Because Since herpesviruses bud through the nuclear membrane, the set of lipids in their supercapsid reflects the composition of the nuclear membrane. The sugars that make up glycoproteins also come from the host cell.
On the inner surface of the supercapsid there is the so-called matrix proteins (M proteins) a structural layer has been formed that facilitates the interaction of the supercapsid with the nucleocapsid, which is extremely important at the final stages of virion self-assembly.
However, the main structural and functional component of the virus is its genome, which determines all the properties of the viral particle, both inside and outside the target cell. The genome contains encrypted information about the morphological, biochemical, pathogenic and antigenic properties of its carrier. The genome of the viral particle is haploid. Nucleic acids are represented by single-stranded RNA molecules or double-stranded DNA molecules. The exceptions are reoviruses, whose genome is formed by two strands of RNA, and parvoviruses, whose genome is presented as a single strand of DNA. Viruses contain only one type of nucleic acid.
Viral DNA are organized as circular covalently linked supercoiled or linear structures with a molecular weight of 1 10 6 to 1 10 8, which is 10 to 100 times less than the molecular weight of bacterial DNA. The genome contains up to several hundred genes. Transcription of viral DNA occurs in the nucleus of the infected cell . Nucleotide sequences occur once, but at the ends of the molecule there are straight and inverted (turned 180 o) repeating nucleotide sequences. This ensures the ability of the DNA molecule to close into a ring. In addition, they are unique markers of viral DNA.
Viral RNAs are represented by single- and double-stranded molecules and in their chemical composition do not differ from RNA of cellular origin. Single-stranded molecules can be segmented, which leads to an increase in the coding capacity of the genome. In addition, they contain helical regions such as a DNA double helix, formed by pairing of complementary nitrogenous bases. Double-stranded RNA can be linear or circular.
Depending on the specific intracellular behavior and functions performed, viral RNAs are divided into groups:
1. Plus strand RNA, having the ability to translate the information encoded in it to the ribosomes of the target cell, i.e. perform the function of mRNA. RNAs of plus-strand viruses have characteristic modified cap-shaped ends necessary for specific recognition of ribosomes. They are called plus strands or positive genome.
2. Minus strand RNA are not capable of translating genetic information directly to ribosomes and cannot function as mRNA. However, they are a template for the synthesis of mRNA. They are called minus strands or negative genome.
3. Double threads one of which functions as –RNA, the other, complementary to it, as +RNA.
Many viral nucleic acids +RNA and DNA-containing viruses are infectious in themselves, because contain all the genetic information necessary for the synthesis of new viral particles. This information is realized after the virion penetrates the sensitive cell. Double-stranded RNAs and most -RNAs cannot exhibit infectious properties.
The interaction of a virus with a target cell is a complex and multi-stage process of coexistence of two forms of living matter – precellular and cellular. Here the entire complex of effects of the viral genome on the genetically encoded biosynthetic processes of the host cell is manifested.
The implementation of the reproductive cycle largely depends on the type of infection of the cell and the nature of the interaction of the virus with the sensitive (possibly infected) cell.
In a virus-infected cell, viruses can remain in various states:
1. reproduction of numerous new virions;
2. the presence of the virus nucleic acid in an integrated state with the cell chromosome in the form of a provirus;
3. existence in the cytoplasm of the cell in the form of circular nucleic acids, reminiscent of bacterial plasmids.
It is these conditions that determine a wide range of disorders caused by the virus: from a pronounced productive infection ending in cell death, to prolonged interaction of the virus with the cell in the form of a latent (latent) infection or malignant transformation of the cell.
Four types of interaction of the virus with a sensitive cell have been identified:
1. Productive type – ends with the formation of a new generation of virions and their release as a result of lysis of infected cells ( cytolytic form), or leaving the cell without destroying it ( noncytolytic form). The non-cytolytic type of interaction most often occurs persistent chronic infections, characterized by the formation of daughter populations of the pathogen after completion of the acute phase of the disease. Cell death is caused by early suppression of the synthesis of cellular proteins, accumulation of toxic and specifically damaging viral components, damage to lysosomes and the release of their enzymes into the cytoplasm;
2. Integrative type , or virogeny – characterized by the incorporation (integration) of viral DNA in the form of a provirus into the cell chromosome and subsequent functioning as its integral part with joint replication. This type of interaction proceeds latent infection, lysogeny of bacteria And viral transformation of cells;
3. Abortive type – does not end with the formation of new virions, since the infectious process in the cell is interrupted at one of the stages. Occurs when a virus interacts with a resting cell, or when a cell is infected with a defective virus.
Both viruses and virions can be defective.
Defective viruses exist as independent species and are functionally inferior, because For their replication, a “helper virus” is needed, i.e. the defect is determined by the inferiority of the genome. Divided into 3 groups:
1. Defective Interfering Particles, which are virions containing only part of the genetic information of the original virus and replicate only with the participation of a related “helper virus”;
2. Satellite viruses They differ from the previous ones in that for their reproduction they require the participation of any “helper virus”, not necessarily related;
3. Integrated Genomes are proviruses, i.e. viral genomes that are integrated into the chromosome of a cell, but have lost the ability to transform into a full-fledged virus;
Defective virionsconstitute a group formed during the formation of large daughter populations, and their defectiveness is determined mainly by morphological inferiority (empty capsids, non-enveloped nucleocapsids, etc.). A special form of defective virions is pseudovirions, having a normal capsid containing part of its own nucleic acid and fragments of the host nucleic acid, or part of the host cell chromosome and part of the nucleic acid of another virus.
The significance of defective viruses is their ability to transfer genetic material from a donor cell to a recipient cell.
4. Virus interference – occurs when a cell is infected by two viruses and does not occur with any combination of pathogens. Interference is realized either due to the induction by one virus of cellular inhibitors that suppress the reproduction of the other, or due to damage to the receptor apparatus or cell metabolism by the first virus, which excludes the possibility of reproduction of the second. Distinguish homologous(related viruses) and heterologous(unrelated viruses) interference.
Based on the nature of the interaction of the virus genome with the genome of cells, they are distinguished autonomous And integration infection. During autonomous infection, the viral genome is not integrated into the cell genome, while during integration infection, the viral genome is integrated into the cellular one.
Productive type of virus-cell interaction , i.e. Viral reproduction is a unique form of expression of foreign (viral) genetic information in the cells of humans, animals, plants and bacteria, which consists in subordinating the cellular matrix-genetic mechanisms of viral information. This is a complex process of interaction between two genomes occurring in 6 stages:
1. adsorption of virions;
2. penetration of the virus into the cell;
3. “undressing” and release of the viral genome;
4. synthesis of viral components;
5. formation of virions;
6. exit of virions from the cell.
First reproduction stage – adsorption, i.e. attachment of the virion to the cell surface. It occurs in two phases. First phase – nonspecific, caused by ionic attraction and other mechanisms of interaction between the virus and the cell. Second phase – highly specific, due to the homology and complementarity of the receptors of sensitive cells and the protein ligands of viruses that recognize them. Viral proteins that recognize and interact are called attachment and are represented by glycoproteins, part of the lipoprotein shell of the capsid or supercapsid of the virus.
Specific cell receptors have a different nature, being proteins, lipids, carbohydrate components of proteins and lipids. One cell can carry from ten to one hundred thousand specific receptors, which allows tens and hundreds of virions to attach to it. The number of infectious viral particles adsorbed on the cell determines the term "multiplicity of infection". However, a cell infected with a virus is in most cases tolerant to reinfection by a homologous virus.
The presence of specific receptors underlies tropism viruses to certain cells, tissues and organs.
Second stage – entry of the virus into the cell can happen in several ways.
1. Receptor-dependent endocytosis occurs as a result of capture and absorption of the virion by a sensitive cell. In this case, the cell membrane with the attached virion invaginates to form an intracellular vacuole (endosome) containing the virus. Next, the lipoprotein shell of the virus merges with the endosome membrane and the virus exits into the cell cytoplasm. Endosomes combine with lysosomes, which destroy the remaining viral components.
2. Viropexis – consists of the fusion of the viral supercapsid with the cellular or nuclear membrane and occurs with the help of a special fusion protein – F protein, which is part of the supercapsid. As a result of viropexis, the capsid ends up inside the cell, and the supercapsid, together with the protein, integrates (embeds itself) into the plasma or nuclear membrane. Inherent only in complex viruses.
3. Phagocytosis – by means of which viruses penetrate into phagocytic cells, which leads to incomplete phagocytosis.
Third stage – "undressing" and releasing the viral genome occurs as a result of deproteinization, modification of the nucleocapsid, removal of surface viral structures and release of an internal component that can cause an infectious process. The first stages of “undressing” begin during the process of penetration into the cell by the fusion of viral and cellular membranes or when the virus exits the endosome into the cytoplasm. Subsequent stages are closely related to their intracellular transport to the sites of deproteinization. Different viruses have their own specialized “undressing” areas. Transport to them is carried out using intracellular membrane vesicles, in which the virus is transferred to ribosomes, the endoplasmic reticulum or the nucleus.
Fourth stage – synthesis of viral components starts at the moment shadow or eclipse phases, which is characterized by the disappearance of the virion. The shadow phase ends after the formation of the constituent components of the virus necessary for the assembly of daughter populations. The virus uses the genetic apparatus of the cell for this, suppressing the synthetic reactions it itself needs. Synthesis of proteins and nucleic acids of the virus, i.e. its reproduction, separated in time and space, occurs in different parts of the cell and is called disjunctive.
In an infected cell, the viral genome encodes the synthesis of two groups of proteins:
- non-structural proteins servicing the intracellular reproduction of the virus at its different stages, which include RNA or DNA polymerases that ensure transcription and replication of the viral genome, regulatory proteins, precursors of viral proteins, enzymes that modify viral proteins;
- structural proteins, included in the virion (genomic, capsid and supercapsid).
Protein synthesis in the cell is carried out in accordance with the processes transcriptions by “rewriting” genetic information from nucleic acid into the nucleotide sequence of messenger RNA (mRNA) and broadcasts(reading) mRNA on ribosomes to form proteins. The term "translation" refers to the mechanisms by which the sequence of nucleic bases of mRNA is translated into a specific sequence of amino acids in the synthesized polypeptide. In this case, discrimination of cellular mRNAs occurs and synthetic processes on ribosomes come under viral control. The mechanisms of information transfer regarding mRNA synthesis are different in different groups of viruses.
Double-stranded DNA containing viruses implement genetic information in the same way as the cellular genome, according to the scheme: genomic DNA of the virus→ mRNA transcription→viral protein translation. At the same time, DNA-containing viruses, the genomes of which are transcribed in the nucleus, use cellular polymerase for this process, and the genomes of which are transcribed in the cytoplasm use their own virus-specific RNA polymerase.
Genome –RNA viruses serves as a template from which mRNA is transcribed with the participation of a virus-specific RNA polymerase. Their protein synthesis occurs according to the following scheme: genomic RNA of the virus→mRNA transcription→viral protein translation.
A special group of RNA-containing retroviruses stands out, which includes human immunodeficiency viruses and oncogenic retroviruses. They have a unique way of transmitting genetic information. The genome of these viruses consists of two identical RNA molecules, i.e. is diploid. Retroviruses contain a special virus-specific enzyme - reverse transcriptase, or revertase, with the help of which the process of reverse transcription is carried out. It consists of the following: complementary single-stranded DNA (cDNA) is synthesized on the genomic RNA matrix. It is copied to form double-stranded complementary DNA, which is integrated into the cellular genome and, within its composition, transcribed into mRNA using cellular DNA-dependent RNA polymerase. The synthesis of proteins of these viruses is carried out according to the following scheme: genomic RNA of the virus→complementary DNA→mRNA transcription→viral protein translation.
Transcription regulation is carried out by cellular and virus-specific mechanisms. It consists of sequentially reading information from the so-called. "early" And "late" genes. The first encodes information for the synthesis of virus-specific transcription and replication enzymes, and the second - for the synthesis of capsid proteins.
Synthesis of viral nucleic acids, i.e. replication of viral genomes, leads to the accumulation in the cell of copies of the original viral genomes, which are used in the assembly of virions. The method of replication depends on the type of virus nucleic acid, the presence of virus-specific and cellular polymerases, and the ability of viruses to induce the formation of polymerases in the cell.
Double-stranded DNA viruses replicate in the usual semi-conservative way: after the DNA strands unwind, new strands are complementarily added to them. Each newly synthesized DNA molecule consists of one parent and one synthesized strand.
Single-stranded DNA viruses During the replication process, cellular DNA polymerases are used to create a double-stranded viral genome, the so-called. replicative form. In this case, a –DNA strand is complementarily synthesized on the original +DNA strand, which serves as a template for the +DNA strand of the new virion.
Single-stranded +RNA viruses induce the synthesis of RNA-dependent RNA polymerase in the cell. With its help, on the basis of the genomic +RNA strand, the –RNA strand is synthesized, and a temporary double RNA is formed, called intermediate replicative link. It consists of a complete +RNA strand and numerous partially completed –RNA strands. When all the –RNA strands are formed, they are used as templates for the synthesis of new +RNA strands.
Single-stranded RNA viruses contain an RNA-dependent RNA polymerase. The genomic –RNA strand is transformed by the viral polymerase into incomplete and complete +RNA strands. Incomplete copies act as mRNA for the synthesis of viral proteins, and complete copies serve as a template for the synthesis of the genomic RNA strand of the offspring.
Double-stranded RNA viruses replicate similarly to single-stranded RNA viruses. The difference is that the +RNA strands formed during transcription function not only as mRNA, but also participate in replication. They are the template for the synthesis of RNA strands. Together they form genomic double-stranded RNA virions.
Diploid +RNA viruses or retroviruses are replicated using viral reverse transcriptase, which synthesizes a –DNA strand on the RNA virus template, from which the +DNA strand is copied to form a double strand of DNA closed in a ring. Next, the double strand of DNA integrates with the cell chromosome, forming a provirus. Numerous virion RNAs are formed as a result of transcription of one of the integrated DNA strands with the participation of cellular DNA-dependent RNA polymerase.
Fifth stage – virion assembly occurs through an orderly self-assembly, when the components of the virion are transported to the site of virus assembly. These are specific areas of the nucleus and cytoplasm called replication complexes. The connection of the virion components is due to the presence of hydrophobic, ionic, hydrogen bonds and stereochemical correspondence.
The formation of viruses is a multi-stage, strictly sequential process, with the formation of intermediate forms that differ from mature virions in the composition of polypeptides. The assembly of simply arranged viruses occurs on replication complexes and consists of the interaction of viral nucleic acids with capsid proteins and the formation of nucleocapsids. In complex viruses, nucleocapsids are first formed on replication complexes, which then interact with modified cell membranes, which are the future lipoprotein shell of the virion. In this case, the assembly of viruses replicating in the nucleus occurs with the participation of the nuclear membrane, and the assembly of viruses whose replication occurs in the cytoplasm is carried out with the participation of the membranes of the endoplasmic reticulum or the cytoplasmic membrane, where glycoproteins and other proteins of the virion envelope are embedded. In some complex RNA viruses, a matrix protein is involved in assembly - M protein– which is located under the cell membrane modified by this protein. Possessing hydrophobic properties, it acts as an intermediary between the nucleocapsid and the supercapsid. Complex viruses include components of the host cell during their formation. When the self-assembly process is disrupted, “defective” virions are formed.
Sixth stage – release of viral particles from the cell completes the process of viral reproduction and occurs in two ways.
Explosive path, when viruses lacking a supercapsid cause cell destruction and enter the extracellular space. A large number of virions emerge from a dead cell at the same time.
Budding or exocytosis , characteristic of complex viruses whose supercapsid is derived from cell membranes. First, the nucleocapsid is transported to cell membranes, into which virus-specific proteins are already embedded. In the area of contact, protrusion of these areas begins to form a kidney. The formed bud is separated from the cell in the form of a complex virion. The process is not lethal to the cell and the cell is able to remain viable for a long time, producing viral progeny.
Budding of viruses formed in the cytoplasm can occur either through the plasma membrane or through the membranes of the endoplasmic reticulum and Golgi apparatus, followed by release to the cell surface.
Viruses that form in the nucleus bud into the perinuclear space through the modified nuclear envelope and are transported to the cell surface as part of cytoplasmic vesicles.
Integrative type of virus-cell interaction (virogeny) is the coexistence of a virus and a cell as a result of the integration of the viral nucleic acid into the host cell chromosome, in which the viral genome replicates and functions as the main part of the cell genome.
This type of interaction is characteristic of moderate DNA-containing bacteriophages, oncogenic viruses and some infectious DNA and RNA viruses.
Integration requires the presence of a circular form of double-stranded viral DNA. Such DNA is attached to cellular DNA at the site of homology and is inserted into a specific region of the chromosome. For RNA viruses, the integration process is more complex and begins with the reverse transcription mechanism. Integration occurs after the formation of a double-stranded DNA transcript and its closure into a ring.
Additional genetic information during virogenesis imparts new properties to the cell, which can cause oncogenic transformation of cells, autoimmune and chronic diseases.
Abortive type of virus-cell interaction does not end with the formation of viral progeny and can occur under the following conditions:
1. infection of a sensitive cell occurs with a defective virus or a defective virion;
2. infection of genetically resistant cells by a virulent virus;
3. infection of a sensitive cell with a virulent virus non-permissive (non-permissive) conditions.
More often, an abortive type of interaction is observed when an insensitive cell is infected with a standard virus. However, the mechanism of genetic resistance is not the same. It may be associated with the absence of specific receptors on the plasma membrane, the inability of this type of cell to initiate translation of viral mRNA, and the absence of specific proteases or nucleases necessary for the synthesis of viral macromolecules.
Changes in the conditions in which viral reproduction occurs can also lead to abortive interaction: an increase in body temperature, a change in pH at the site of inflammation, the introduction of antiviral drugs, etc. However, when non-permissive conditions are eliminated, the abortive type of interaction becomes productive with all the ensuing consequences.
Interfering interaction is determined by the state of immunity to secondary infection of a cell already infected with the virus.
Heterologous interference occurs when infection with one virus completely blocks the ability of a second virus to replicate within one cell. One of the mechanisms is associated with inhibition of the adsorption of another virus by blocking or destroying specific receptors. Another mechanism involves inhibition of mRNA translation by any heterologous mRNA in the infected cell.
Homologous interference typical of many defective viruses, especially those that are repeatedly passaged in vitro and with a high multiplicity of infection. Their reproduction is possible only when the cell is infected with a normal virus. Sometimes a defective virus can interfere with the reproductive cycle of a normal virus and produce defective interfering viral particles (VIPs). DI particles contain only part of the genome of a normal virus. By the nature of the defect, DI particles are deletion and can be considered lethal mutants. The main property of DI particles is the ability to interfere with a normal homologous virus and can even play the role of assistants during replication. The ability to adsorb and penetrate into the cell is associated with the normal structure of the capsid. The release and expression of a defective nucleic acid leads to various biological effects: it inhibits synthetic processes in the cell and, due to homologous interference, inhibits the synthesis and transformation of proteins of normal viruses. Circulation of DI particles and co-infection with a normal homologous virus causes the appearance of sluggish, long-term forms of disease, which is associated with the ability of DI particles to replicate much faster due to the simplicity of the genome, while the defective population has a noticeable decrease in the severity of the cytopathic effect characteristic of a normal virus.
The process of interaction of the virus with the body in most cases is cytospecific and is determined by the ability of the pathogen to reproduce in certain tissues. However, some viruses have a wider range of tropism and reproduce in a wide variety of cells and organs.
The specificity factors of the virus, responsible for its tropism and the diversity of the cells affected, include the number of specific receptors (both in the virion and in the cell) that ensure full interaction of the virus with the cell. The number of such receptors is usually limited.
In some cases, the very physiological specificity of cells, and therefore their bimolecular organization, contributes to the manifestation of the virulence of the pathogen. For example, the G-protein of the rabies virus envelope has a high affinity for acetylcholine receptors of neurons, which ensures its ability to penetrate cells of nervous tissue. It should be noted that neurotropic viruses cause especially severe diseases, because nerve cells do not regenerate. Moreover, the reproduction of the pathogen makes them targets for cytotoxic immune reactions.
Quite often, the virulence of viruses increases due to mutations. Of particular importance in this case is the ability of viruses to reverse gene mutation (reversion). Genes encoding protein structure can restore their structure and transform previously avirulent virus strains into virulent ones.
Equally important are characteristics of a susceptible microorganism.
Age is about
Microbiology: lecture notes Ksenia Viktorovna Tkachenko
1. Morphology and structure of viruses
Viruses are microorganisms that make up the kingdom Vira.
Features:
2) do not have their own protein-synthesizing and energy systems;
3) do not have a cellular organization;
4) have a disjunctive (separated) method of reproduction (synthesis of proteins and nucleic acids occurs in different places and at different times);
6) viruses pass through bacterial filters.
Viruses can exist in two forms: extracellular (virion) and intracellular (virus).
The shape of virions can be:
1) round;
2) rod-shaped;
3) in the form of regular polygons;
4) thread-like, etc.
Their sizes range from 15–18 to 300–400 nm.
In the center of the virion is a viral nucleic acid, covered with a protein shell - a capsid, which has a strictly ordered structure. The capsid shell is made up of capsomeres. Nucleic acid and the capsid shell make up the nucleocapsid.
The nucleocapsid of complexly organized virions is covered with an outer shell - a supercapsid, which can include many functionally different lipid, protein, and carbohydrate structures.
The structure of DNA and RNA viruses is not fundamentally different from the NK of other microorganisms. Some viruses contain uracil in their DNA.
DNA can be:
1) double-stranded;
2) single-chain;
3) ring;
4) double-stranded, but with one shorter chain;
5) double-chain, but with one continuous and the other fragmented chains.
RNA can be:
1) single thread;
2) linear double-stranded;
3) linear fragmented;
4) ring;
Viral proteins are divided into:
1) genomic – nucleoproteins. Provide replication of viral nucleic acids and viral reproduction processes. These are enzymes, due to which the number of copies of the parent molecule increases, or proteins, with the help of which molecules are synthesized on a nucleic acid matrix that ensure the implementation of genetic information;
2) capsid shell proteins are simple proteins with the ability to self-assemble. They form geometrically regular structures, in which several types of symmetry are distinguished: spiral, cubic (they form regular polygons, the number of faces is strictly constant) or mixed;
3) supercapsid shell proteins are complex proteins with diverse functions. Due to them, the interaction of viruses with a sensitive cell occurs. Perform protective and receptor functions.
Among the proteins of the supercapsid shell are:
a) anchor proteins (one end of them is located on the surface, and the other goes deep; they ensure contact of the virion with the cell);
b) enzymes (can destroy membranes);
c) hemagglutinins (cause hemagglutination);
d) elements of the host cell.
From the book On the Origin of Species by Natural Selection or the Preservation of Favored Breeds in the Struggle for Life by Darwin CharlesMorphology. We have seen that members of the same class, regardless of their mode of life, are similar to each other in the general plan of organization. This similarity is often expressed by the term "unity of type" or by indicating that certain parts and organs in different species are of the same type.
From the book Fundamentals of Neurophysiology author Shulgovsky Valery ViktorovichGLIA - MORPHOLOGY AND FUNCTION The human brain consists of hundreds of billions of cells, with nerve cells (neurons) not making up the majority. Most of the volume of nervous tissue (up to 9/10 in some areas of the brain) is occupied by glial cells (from the Greek: glue). The fact is that
From the book Microbiology: lecture notes author Tkachenko Ksenia ViktorovnaLECTURE No. 2. Morphology and ultrastructure of bacteria 1. Features of the structure of a bacterial cell. Main organelles and their functions Differences between bacteria and other cells1. Bacteria are prokaryotes, that is, they do not have a separate nucleus.2. In the cell wall of bacteria
From the book Microbiology author Tkachenko Ksenia Viktorovna3. Cultivation of viruses The main methods of cultivating viruses: 1) biological - infection of laboratory animals. When an animal becomes infected with a virus, it becomes sick. If the disease does not develop, then pathological changes can be detected at autopsy. In animals
From the book General Ecology author Chernova Nina Mikhailovna1. Morphology and cultural properties The pathogen belongs to the genus Carinobakterium, species C. difteria. These are thin rods, straight or slightly curved, gram-positive. They are characterized by pronounced polymorphism. At the ends there are club-shaped thickenings - metachromatic grains of volutin.
From the book Biology [Complete reference book for preparing for the Unified State Exam] author Lerner Georgy Isaakovich1. Morphology and cultural properties The pathogen belongs to the genus Mycobakterium, species M. tuberculesis. These are thin rods, slightly curved, do not form spores or capsules. The cell wall is surrounded by a layer of glycopeptides called mycosides (microcapsules). Tuberculosis bacillus
From the book Journey to the Land of Microbes author Betina Vladimir4. Morphology of bacteria, main organs The sizes of bacteria range from 0.3-0.5 to 5-10 microns. Based on the shape of the cells, bacteria are divided into cocci, rods and convoluted. In a bacterial cell there are: 1) main organelles: (nucleoid, cytoplasm , ribosomes, cytoplasmic
From the book The Newest Book of Facts. Volume 1. Astronomy and astrophysics. Geography and other earth sciences. Biology and medicine author Kondrashov Anatoly Pavlovich5. Morphology of bacteria, additional organelles Villi (pili, fimbriae) are thin protein outgrowths on the surface of the cell wall. Comon pili are responsible for the adhesion of bacteria to the surface of cells of the macroorganism. They are characteristic of gram-positive bacteria.
From the book Clematis author Beskaravainaya Margarita Alekseevna10. Morphology of viruses, types of interaction of a virus with a cell Viruses are microorganisms that make up the kingdom Vira. Viruses can exist in two forms: extracellular (virion) and intracellular (virus). In shape, virions can be: round, rod-shaped,
From the book Logic of Chance [On the nature and origin of biological evolution] author Kunin Evgeniy ViktorovichChapter 6. ADAPTIVE MORPHOLOGY OF ORGANISMS Among the adaptations of animals and plants to the environment, morphological adaptations play an important role, i.e., such features of the external structure that contribute to the survival and successful functioning of organisms in normal conditions.
From the author's book From the author's bookInfectious RNA and reconstruction of viruses Evidence that viral RNA is genetic material was provided to us by the same TMV. First of all, scientists managed to change TMV particles by eliminating the protein component from their composition. In this state, viruses
From the author's bookThe Threat of Viruses One of the books about viruses is very aptly titled “Viruses - the Enemies of Life.” And not only influenza viruses, but also other viruses that infect humans, are responsible for tens of thousands, and perhaps millions of lives. Rubella should be considered an unsafe disease. This
From the author's book From the author's bookMorphology and biology of clematis Clematis? perennial, overwhelmingly deciduous, rarely evergreen, plants. Root system. Mature clematis have two main types of root systems: taproot and fibrous. With limited watering (in the south)
From the author's bookChapter 10 The world of viruses and its evolution Trans. G. Janus viruses were discovered as something completely unremarkable, namely an unusual variety of infectious agents, and possibly a special kind of toxin that causes plant diseases, for example, tobacco mosaic. Since these agents
Viruses are microorganisms that make up the kingdom Vira.
Features:
2) do not have their own protein-synthesizing and energy systems;
3) do not have a cellular organization;
4) have a disjunctive (separated) method of reproduction (synthesis of proteins and nucleic acids occurs in different places and at different times);
6) viruses pass through bacterial filters.
Viruses can exist in two forms: extracellular (virion) and intracellular (virus).
The shape of virions can be:
1) round;
2) rod-shaped;
3) in the form of regular polygons;
4) thread-like, etc.
Their sizes range from 15–18 to 300–400 nm.
In the center of the virion is a viral nucleic acid, covered with a protein shell - a capsid, which has a strictly ordered structure. The capsid shell is made up of capsomeres. Nucleic acid and the capsid shell make up the nucleocapsid.
The nucleocapsid of complexly organized virions is covered with an outer shell - a supercapsid, which can include many functionally different lipid, protein, and carbohydrate structures.
The structure of DNA and RNA viruses is not fundamentally different from the NK of other microorganisms. Some viruses contain uracil in their DNA.
DNA can be:
1) double-stranded;
2) single-chain;
3) ring;
4) double-stranded, but with one shorter chain;
5) double-chain, but with one continuous and the other fragmented chains.
RNA can be:
1) single thread;
2) linear double-stranded;
3) linear fragmented;
4) ring;
Viral proteins are divided into:
1) genomic – nucleoproteins. Provide replication of viral nucleic acids and viral reproduction processes. These are enzymes, due to which the number of copies of the parent molecule increases, or proteins, with the help of which molecules are synthesized on a nucleic acid matrix that ensure the implementation of genetic information;
2) capsid shell proteins are simple proteins with the ability to self-assemble. They form geometrically regular structures, in which several types of symmetry are distinguished: spiral, cubic (they form regular polygons, the number of faces is strictly constant) or mixed;
3) supercapsid shell proteins are complex proteins with diverse functions. Due to them, the interaction of viruses with a sensitive cell occurs. Perform protective and receptor functions.
Among the proteins of the supercapsid shell are:
a) anchor proteins (one end of them is located on the surface, and the other goes deep; they ensure contact of the virion with the cell);
b) enzymes (can destroy membranes);
c) hemagglutinins (cause hemagglutination);
d) elements of the host cell.
2. Interaction of viruses with the host cell
The interaction takes place in a single biological system at the genetic level.
There are four types of interaction:
1) productive viral infection (interaction as a result of which virus reproduction occurs and cells die);
2) abortive viral infection (an interaction in which virus reproduction does not occur, and the cell restores the impaired function);
3) latent viral infection (the virus reproduces, but the cell retains its functional activity);
4) virus-induced transformation (an interaction in which a cell infected with a virus acquires new properties that were not previously inherent to it).
After adsorption, virions penetrate inside by endocytosis (viropexis) or as a result of the fusion of viral and cellular membranes. The resulting vacuoles, containing entire virions or their internal components, enter lysosomes, in which deproteinization occurs, i.e., “undressing” of the virus, as a result of which the viral proteins are destroyed. Viral nucleic acids freed from proteins penetrate through cellular channels into the cell nucleus or remain in the cytoplasm.
Nucleic acids of viruses implement the genetic program for the creation of viral offspring and determine the hereditary properties of viruses. With the help of special enzymes (polymerases), copies are made from the parent nucleic acid (replication occurs), and messenger RNAs are also synthesized, which connect to ribosomes and carry out the synthesis of daughter viral proteins (translation).
After a sufficient number of virus components have accumulated in the infected cell, the assembly of progeny virions begins. This process usually occurs near cell membranes, which sometimes take a direct part in it. Newly formed virions often contain substances characteristic of the cell in which the virus multiplies. In such cases, the final stage of virion formation is enveloping them in a layer of cell membrane.
The last stage of interaction between viruses and cells is the exit or release of daughter viral particles from the cell. Simple viruses lacking a supercapsid cause cell destruction and enter the intercellular space. Other viruses that have a lipoprotein envelope exit the cell by budding. In this case, the cell remains viable for a long time. In some cases, viruses accumulate in the cytoplasm or nucleus of infected cells, forming crystal-like clusters - inclusion bodies.
The morphology and structure of viruses is studied using an electron microscope, since their sizes are small and comparable to the thickness of the bacterial shell.
The shape of virions can be different: rod-shaped (tobacco mosaic virus), bullet-shaped (rabies virus), spherical (poliomyelitis viruses, HIV), sperm-shaped (many bacteriophages) (Fig. 8).
Rice. 8. Forms of virions:
1 smallpox virus; 2 herpes virus; 3 adenovirus; 4 papovavirus; 5 hepadnavirus; 6 paramyxovirus; 7 influenza virus; 8 coronavirus; 9 arenavirus; 10 retrovirus;
The sizes of viruses are determined using electron microscopy, ultrafiltration through filters with a known pore diameter, and ultracentrifugation. Some of the smallest viruses are polio and foot-and-mouth disease viruses (about 20 nm), circoviruses (16 nm), and the largest variola virus (about 350 nm). Viruses have a unique genome because they contain either DNA or RNA. Therefore, a distinction is made between DNA-containing and RNA-containing viruses. They are usually haploid, meaning they have one set of genes. The genome of viruses is represented by various types of nucleic acids: double-stranded, single-stranded, linear, circular, fragmented.
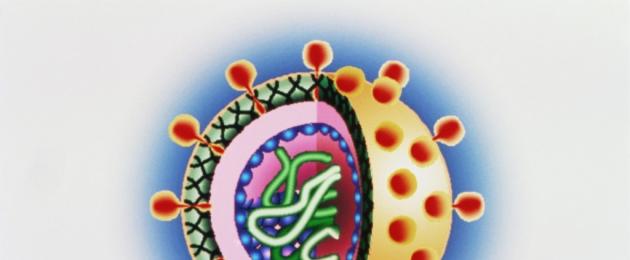
There are simple viruses (for example, polio virus) and complex viruses (for example, influenza viruses, measles viruses). In simple viruses, the nucleic acid is associated with a protein shell called a capsid (from the Latin capsa case). The capsid consists of repeating morphological subunits of capsomeres. Nucleic acid and capsid interact with each other to form a nucleocapsid. In complex viruses, the capsid is surrounded by an additional lipoprotein shell, a supercapsid (a derivative of the membrane structures of the host cell), which has “spikes”. The capsid and supercapsid protect virions from environmental influences, determine selective interaction (adsorption) with cells, and determine the antigenic and immunogenic properties of virions. The internal structures of viruses are called the core.
Virions are characterized by spiral, cubic and complex types of capsid symmetry. The helical type of symmetry is due to the helical structure of the nucleocapsid, the cubic formation of an isometric hollow body from the capsid containing the viral nucleic acid.
In addition to ordinary viruses, so-called non-canonical viruses are also known: prions, protein infectious particles that have the form of fibrils measuring 10-20 x 100-200 nm. Prions, apparently, are both inducers and products of an autonomous gene in humans or animals and cause encephalopathy in them under conditions of slow viral infection (Creutzfeldt-Jakob disease, kuru, etc.). Other unusual agents closely related to viruses are viroids, small circular, supercoiled RNA molecules that do not contain protein and cause diseases in plants.
The scene is the laboratory of the Nikitsky Botanical Garden at the Russian Academy of Sciences, where biologist Dmitry Iosifovich Ivanovsky (1864-1920) is studying the mysterious mosaic disease of tobacco. The causative agent of the disease in a plant passes through the smallest bacterial filters, does not grow on and does not produce symptoms when healthy plants are infected with filtrates from diseased plants.
It was then, in 1892, that the scientist concluded that these were not bacteria. And he calls the pathogen viruses (from the Latin virus - poison). Dmitry Ivanovsky tried to see viruses all his life, but we saw the morphology of viruses in the 30s of the 20th century, when electron microscopes were invented.
But this date is considered the beginning of the science of virology, and Dmitry Ivanovsky is its founder.

Amazing kingdom
The distinctive features of viruses are as follows:

Part of the organic world of the planet
To date, more than 6 thousand viruses have already been described, but it is estimated that there are more than one hundred million of them. This is the most numerous biological form on the planet, and it is represented in all ecosystems (universal (ubiquitous) distribution).
Their appearance on the planet is still unclear. One thing is known - when the first cellular life forms appeared, viruses already existed.

Alive and not alive
These amazing organisms have two forms of their existence, which are significantly different from each other.
Virion is essentially a non-living part of life. And the genome of the virus in the cell is its living component, because this is where viruses reproduce.

Morphology and ultrastructure of viruses
In this context, we are talking about the virion - the extracellular form.
The size of virions is measured in nanometers - 10 -9 meters. Influenza viruses have an average size of 80-120 nanometers, and the smallpox virus is a giant with a size of 400 nanometers.
The structure and morphology of viruses is similar to astronauts. Inside the capsid (a protein shell, sometimes containing fats and carbohydrates), like in a “space suit,” there is the most valuable part - nucleic acids, the genome of the virus. Moreover, this “cosmonaut” is presented in a minimal quantity - only directly hereditary material and a minimum of enzymes for its replication (copying).
Externally, the “spacesuit” can be rod-shaped, spherical, bullet-shaped, in the shape of a complex icosahedron, or even irregular in shape. This depends on the presence in the capsid of specific proteins that are responsible for the penetration of the virus into the cell.

How does a pathogen enter the host?
There are many methods of penetration, but the most common is airborne. Myriads of tiny particles are thrown into space not only when coughing or sneezing, but also simply when breathing.
Another way virions enter the body is contagious (direct physical contact). This method is characteristic of a fairly small group of pathogens; this is how herpes, sexually transmitted infections, and AIDS are transmitted.
The method of infection through a vector, which can be various groups of organisms, is quite complex. The vector, having received the pathogen from the reservoir of infection, becomes a place where viruses can multiply or undergo development stages. The rabies virus is just such a pathogen.

What happens in the host's body
With the help of external capsid proteins, the virus attaches to the cell membrane and penetrates through endocytosis. They enter lysosomes, where, under the action of enzymes, they get rid of the “spacesuit”. And the nucleic acids of the pathogen enter the nucleus or remain in the cytoplasm.
The pathogen's nucleic acids are integrated into the host's nucleic acid chains, and the replication (copying) reaction of hereditary information is triggered. When a sufficient number of viral particles have accumulated in the cell, the virions use the energetic and plastic mechanisms and resources of the host.
The last stage is the release of virions from the cell. Some viruses lead to complete destruction of cells and enter the intercellular space, others enter it through exocytosis or budding.

Pathogen Strategies
The structure and morphology of viruses leads to the complete dependence of the pathogen on the energy and protein-synthesizing potential of the cell; the only condition is that it replicates its nucleic acids according to its own schedule. This interaction is called productive (natural for the virus, but not for the cell). Having exhausted the cell's supply, the virus leads to its death.
Another type of interaction is conciliatory. In this case, the viral genome, integrated into the host genome, is replicated covalently with the cell’s own nucleic acids. And then the development of the scenario can go in two directions. The virus behaves quietly and does not manifest itself. Young virions leave the cell only under certain conditions. Either the pathogen's genes are constantly working, producing a large number of young generations, but the cell does not die, and they leave it through exocytosis.
Complexities of taxonomy
The classification and morphology of viruses is different in a variety of sources. In this case, the following characteristics are used to classify them:
- Type of nucleic acid (RNA-containing and DNA-containing) and method of its replication. The most common classification of viruses, proposed by American virologist David Baltimore in 1971.
- Morphology and structure of the virus (single-stranded, double-stranded, linear, circular, fragmented, non-fragmented).
- Dimensions, type of symmetry, number of capsomeres.
- Presence of a supercapsid (outer shell).
- Antigenic properties.
- Type of genetic interaction.
- Circle of potential owners.
- Localization in the host cell - in the nucleus or in the cytoplasm.
It is the choice of the main criterion and the morphology of viruses that in microbiology determines the various approaches to the classification of viruses. It's quite difficult. The difficulty is that we begin to study the morphology and structure of the virus only when they lead to pathological processes.

Picky and not very
Based on host choice, these pathogens are extremely diverse in their preferences. Some attack exclusively one biological species - they have a very strict “registration”. For example, it eats influenza viruses from cats, seagulls, and pigs, which are completely safe for other animals. Sometimes the specialization is surprising - the bacteriophage P-17 virus infects only male individuals of one type of E. coli.
Other viruses behave completely differently. For example, bullet-shaped viruses, whose morphology is similar to a bullet, cause completely different diseases and at the same time their host range is extremely wide. These viruses include the rabies virus, which affects all mammals, or the vesicular stomatitis virus (transmitted, by the way, by insects).
- In contact with 0
- Google+ 0
- OK 0
- Facebook 0