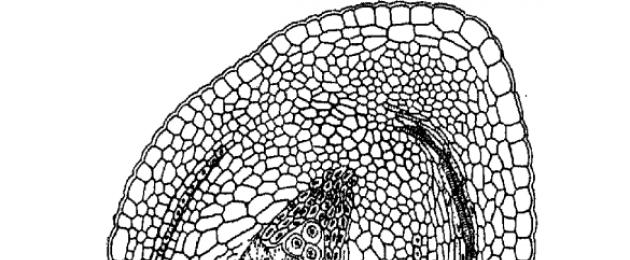
Before moving on to the structure of the ovule, let us consider its main morphological types (Fig. 25), established by the French botanist C. Mirbel in 1829.
In many flowering plants, the micropyle of the ovule is on the same axis with the achene (funiculus) and, thus, the ovule is located perpendicular to the placenta. Such a radially symmetrical ovule was called orthotropic (from the Greek orthos - straight, regular and tropos - turn, direction) or straight. (Fig. 25). It is often called atropic (from the Greek a - particle of negation). Orthotropic ovules are characteristic of Chloranthaceae, Savuraceae, Pepperaceae, Hornwort, Sycamore, Myricaceae, Walnutaceae, Vallisneria, Eelgrass, Pondweed, Xyrisaceae, Eriocaulaceae, Restiaceae and some other genera and families.
, 
Much more common are ovules that are bent downwards (towards the placenta) by 180° and, as a result, adhere to their side to the elongated peduncle. Therefore, their micropyle faces the placenta and is located near the base of the ovule. Such ovules are called anatropic (from the Greek anatrope - inverted, inverted) or inverted (Fig. 25, 26). Anatropic ovules are characteristic of most flowering plants, both dicotyledons and monocotyledons. The famous Czech botanist F. Netolitsky (1926) came to the conclusion that the anatropic type is the original one, and the orthotropic one is the derivative one, which is quite consistent with modern ideas.
Along with anatropic and orthotropic ovules, there are several more “architectural” types, which, however, are less common. First of all, we should mention hemitropic (from the Greek hemi - half, or, more precisely, “half”), or half-inverted, ovules, rotated only 90° relative to the placenta. They are characteristic, for example, of casuarina, some primroses, some borage, Lamiaceae, Solanaceae and Norichaceae, Malpighiaceae, some lilies and a number of other plants. Hemitropic ovules are an intermediate type between anatropic and orthotropic (Fig. 25).
In cases where the ovule, due to stronger growth of one side, becomes curved, it is called campylotropic (from the Greek kampylos - curved) or semi-bent. In contrast to the anatropic and hemitropic types, the integument and even the megasporangium (nucellus) are curved here. This type of ovule is characteristic of many representatives of the order Cloveaceae, capers, some legumes, Philesiaceae, some cereals, and a number of other groups. In most cases, campylotropic ovules arise from anatropic ones, but it is possible that sometimes they can also arise from orthotropic ones.
In some cases, the ovule is bent in the middle in such a way that in a longitudinal section its megasporangium has a horseshoe shape. This is an amphitropic (from the Greek amphi - both) ovule. It is found in Leitneria, Crossosoma, some acanthaceae, and a few other plants (Fig. 27).

Between all these types there are intermediate forms, and therefore it is very often difficult to determine which type of ovule a particular plant has. Nevertheless, the identification of these types turned out to be very useful, since they have a certain systematic significance.
Questions for the colloquium on the topic
for 2nd year biology students
What is an ovule? List its components in Gymnosperms.
Ovule, or ovule(lat. ovulum) - the formation in seed plants from which (usually after fertilization) the seed develops. It is the female sporangium (megasporangium) of seed plants. In gymnosperms - on the surface of the seed scales in female cones, located openly in the axil of the megasporophyll. In the central part of the ovule (nucellus), four megaspores are formed as a result of meiosis of the spore mother cell, then three of them die, and a female gametophyte is formed from one megaspore. In gymnosperms, it is sometimes called endosperm because it stores nutrients in the mature seed.
S. consists of a nucellus (central part containing a megasporocyte), one or two integuments (integuments) and an achene (funicular). At the top of the S. the integument usually does not close, leaving a narrow opening - the micropyle.
What is nucellus? How does the formation and development of the ovule occur?
Nutsellus(from Latin nucella - nut), the central part (core) of the ovule of seed plants; homologous megasporangia ferns. Includes nutritional tissue and a membrane-covered embryo sac with a small opening called micropylem.

Theories of the origin of the ovule integument.
Telomnaya
The integument is the result of the fusion of peripheral initially vegetative bodies around one spore-bearing body. This theory is consistent with paleobotanical findings that relate to proto-gymnosperms and the extinct gymnosperms - seed ferns.
Synangial
According to this hypothesis, first put forward by the English paleobotanist Margarita Benson (1908), the integument is a ring of sterilized, fused and fused sporangia surrounding the central functioning megasporangium, and the micropyle corresponds to the original gap between the tips of the sporangia. In other words, the ovule is actually a synangium in which all sporangia except one have been sterilized and formed the integument of a single, fertile sporangium. A good confirmation of the “synangial” hypothesis is the primitive ovules of seed ferns, which often retain very clear traces of their synangial origin. The ovules of a number of seed ferns had segmented integuments with a vascular bundle in each segment (chamber).
Micropylar and chalazal poles of the ovule
In the seed primordium, the chalaza is located opposite the micropyle, opening the integument. The chalaza is the tissue where the integument and nucellus are connected.
The two poles of the embryo correspond to the root and shoot poles.
In the embryo: hypocotyl and cotyledons
Features of the ovule as a modified megasporangium of seed plants in comparison with the megasporangium of higher spore plants.
Seed is a new type of diaspore.
Megasporogenesis process
There are 2 ovules on the scales of female cones. The ovule consists of a nucellus (nucleus) and integument (cover). A small hole remains at the top - the micropyle (pollen passage). In late spring, the scales of the female cone open and pollen falls through the micropyle inside onto the nucellus. After this, the seed scales are compressed, forming protection for the ovules. During pollination, there are still no male gametes in the dust grain, and the prothallus with archegonia is not yet developed in the ovule. A month after pollination, one of the nucellus cells begins to divide by meiosis. As a result, 4 haploid (n) cells are formed - megaspores. 3 die, and 1 turns into a shoot (n). The female gametophyte is a colorless multicellular thallus. ^ There are many storage substances in the tissue. 14-15 months after pollination, two archegonia are formed on the gametophyte. The archegonium consists of a large egg cell with a large nucleus, above which lies an early disappearing ventral tubule cell, and a neck of eight small cells. At this time, the pollen tube grows very slowly inside the nucellus. After the pollen tube with two sperm reaches the archegonium, one of the sperm fuses with the egg and the other dies. An embryo develops from a zygote. The ovule turns into a seed. The vegetative body of the female gametophyte becomes the primary endosperm (n). A filmy peel is formed from the nuceus, and a woody peel is formed from the integument. Thus, parts of 3 generations can be distinguished in the seed:
Woody and filmy peel – old sporophyte (2n)
Endosperm – gametophyte (n)
Embryo – new sporophyte (2n)
Typical development of the female gametophyte of Gymnosperms

What structures are homologous to the anthers and pollen grains of Gymnosperms?
Anther – microsporangium, speck of dust – pollen grain

10. 2 variants of the structure of a mature male gametophyte, its cellular elements
11. The further fate of the male gametophyte of Gymnosperms.
12. Does a pollen grain always leave the anther mature? Which option is more progressive?
13. Pollination (definition)
In plants, pollen is transferred from the anthers to the stigma (in flowering plants) or to the ovule (in gymnosperms). After O., a pollen tube develops from a speck of dust, the edge grows towards the ovary and is delivered by the husband. sex cells - sperm - to the egg located in the ovule, where fertilization and development of the embryo occurs.
14. Fertilization of the cycad type, its features
After pollination, the ovules begin to enlarge and soon reach the size of a seed, although fertilization has not yet occurred in them. This period, from pollination to fertilization, is very long and usually takes six months (for example, in the cycad, pollination occurs in December - January, and fertilization in May - June).

Microspores that enter the pollen chamber with a drop of pollinating fluid germinate. At the same time, the exine bursts, and the haustorium cell grows through the gap, stretching the intine (Fig. 168, 13). It penetrates the wall of the pollen chamber and sucks nutrients from the nucellus tissue (Fig. 168, 13). At this time, the generative cell divides into two, and one of the resulting cells - spermatogenic - begins to grow rapidly. In it, male gametes - spermatozoa - are formed, not immediately, but after several months (Fig. 168, 15).
By the time of fertilization, the expanded spermatogenic cell is in close proximity to the entrance to the archegonia. The sperm released from it only have to “swim” in the liquid that was poured out with them from the spermatogenic cell, only a short distance to the archegonium, in which the contents of the sperm merge with the egg (Fig. 168, 17).
Thus, in cycads, two mechanisms are combined in a single process, one of which - the formation of a motile sperm - is characteristic of distant ancestors fertilized with the help of water, and the second - the formation of a pollen tube (an expanding spermatogenic cell) - is typical of all others standing higher on the “evolutionary ladder” » seed plants.
15. Siphonogamy
(pine type fertilization) – fertilization using a pollen tube; the sexual process takes place inside the ovules and does not depend on the presence of moisture. The function of delivering male gametes is carried out by special cells.
16. How are these fertilization options fundamentally different in Gymnosperms?
17. What cells (or their features) in the first and second cases can be called atavistic?
The seed develops on the surface of the seed scale. It is a multicellular structure that combines storage tissue - endosperm, embryo and special protective cover (seed coat). Before fertilization, the central part of the ovule contains a nucellus, which is gradually replaced by the endosperm. The endosperm is haploid and is formed from the tissues of the female gametophyte.
U cycads And ginkgo outer layer of the seed coat ( sarcotesta) soft and fleshy, middle layer ( sclerotest) is hard, and the inner layer (endotesta) is filmy at the time the seed ripens. The seeds are dispersed by various animals, which eat the sarcotesta without damaging the sclerotesta.
U yew And podocarpus the seeds are surrounded by fleshy arillus- highly modified scales of a female cone. The succulent and brightly colored aryllus attracts birds, which spread the seeds of these conifers. Arilluses of many species of podocarpus are also edible for humans.
19. From what structures of the ovule do the corresponding parts of the seed develop?
20. The structures of which stages of the life cycle does a seed include?
22. Compare the structure of ginkgo and pine seeds. What are the signs of the primitiveness of the first?
The developed pendulum characteristic of pines degenerates by the time the embryo is fully developed. A pine seed consists of an embryo, a seed coat and a megagametophyte, which is a reserve of nutrients.
In cycads and ginkgo, the outer layer of the seed coat (sarcotesta) is soft and fleshy, the middle layer (sclerotesta) is hard, and the inner layer (endotesta) is filmy at the time the seed ripens.
|
Ginkgo ovules are platyspermic |
||||
|
Additional structures are grouped under the term angiospermization |
||||
|
The pollen chamber is unitegmal |
||||
|
Fertilization after abscission |
||||
23. What is the primary endosperm of a seed? What does its name come from?
The primary endosperm of gymnosperms is formed BEFORE FERTILIZATION from a megaspore and corresponds to the female gametophyte. The endosperm cells of gymnosperms are initially haploid, then, as a result of nuclear fusion, they become polyploid.
Secondary endosperm- tissue formed in the seeds of most flowering plants DURING FERTILIZATION.
24. Evolutionary advantages of seed propagation.
Seeds are more viable, thanks to animals, wind, and water they can be transported over long distances. There is a supply of nutrients, the embryo is protected by the seed coats. Reproduction is not associated with water.
Placentation and its types
Ovary and its types
Structure and types of ovule
General characteristics of the gynoecium. Meaning and structure
The gynoecium is a collection of carpels (carpels). An ovule develops on the carpel, inside which an embryo sac is formed - the female gametophyte, therefore the gynoecium is the female organ of sexual reproduction. Usually the carpels grow together at their edges, forming a closed container - a pistil. The fruit develops from the pistil after fertilization.
For plant reproduction, the gynoecium is of primary importance, because, after fertilization, it grows into a fruit with seeds. In accordance with this significance, different parts of it have different degrees of importance for plants. Its most important part is the ovule, which turns into a seed. Therefore, the gynoecium without ovules loses its biological significance and is considered underdeveloped.
There are no normal plants without ovules; however, there are plants whose gynoecium is represented only by an ovule. Next in importance comes the stigma, that is, the organ that receives pollen. There is no true stigma only in gymnosperms, where it is replaced, however, by loose tissue that develops at the opening of the ovule (micropylar). Less important is the style, which many plants do not have at all (many ranunculaceae, barberry, etc.). Only gymnosperms do not have a true ovary.
The pistil consists of an ovary, a style and a stigma. The same parts are also distinguished in carpels. The ovary carries the ovules, and acts as a moist chamber, protecting the ovules from drying out and being eaten by insects and partly from temperature fluctuations. Seeds develop from the ovules after fertilization. The stigma of the pistil receives pollen, so often the surface of the stigma is uneven, lumpy and covered with a sticky liquid. The style lifts the stigma upward. Sometimes there is no style, and sessile stigmas are formed.
The position of the ovary in the flower depends on the degree of fusion with the receptacle and perianth. There are superior, inferior and semi-inferior ovaries.
The superior ovary is located freely on the receptacle, without merging with other parts of the flower. In this case, parts of the perianth are located under the ovary, so these flowers are also called subpistillate. When the ovary completely fuses with the receptacle and the bases of the petals and stamens, the lower ovary is formed. The flower stalks are attached to the receptacle above the ovary, forming a suprapistal flower. In the semiinferior ovary, only its lower part fuses with the receptacle.
It is assumed that the inferior ovary is more reliably protected from external adverse conditions.
The gynoecium of different plants is extremely diverse not only in its composition, number of parts and the above circumstances, but also in the shape and relative sizes of its parts. In gymnosperms, it consists of two or more ovules that are not closed into an ovary; instead of an ovary, they often have a scale-like leaf, at the base of which they sit (pines, fir, spruce, etc.). Other flowering plants always have a more or less closed ovary, which is why they are called angiosperms. In addition, there are plants in which the entire flower consists of only the gynoecium, even without any covering (willow).
Currently, there are three types of gynoecium:
1) Apocarpous gynoecium - consists of many independent carpels, characterized by marginal placentation.
2) Monocarpous gynoecium - consists of a single pistil with marginal placentation.
3) Coenocarpous gynoecium - consists of several fused pistils. There are three varieties of coenocarpous genicea:
a) Syncarpous - several carpels fused with walls, central-angular placentation.
b) Paracarpous - the same, but the walls of the carpels have not been preserved. Placentation is wall.
c) Lysicarpous - differs from paracarpous by columnar (central) placentation.
The gynoecium can be simple or complex. A compound or apocarpous gynoecium consists of several unfused carpels (raspberry, buttercup).
A simple gynoecium can contain either one carpel, then it is called monocarpous, or one pistil from many fused carpels - this is a coenocarpous gynoecium. There are three subtypes of coenocarpous gynoecium based on the method of carpel fusion: syncarpous, paracarpous and lysicarpous. In a syncarpous gynoecium, the carpels grow together with their lateral surfaces, forming a multilocular ovary, and the ovules are located closer to the center of this ovary (apple tree, cotton). The paracarpous gynoecium is formed by carpels with fused edges, so the ovary is unilocular (grapes, gooseberries). The lysicarpous gynoecium develops from the syncarpous one, but at the same time the walls inside the ovary dissolve and the ovules are located on a column in the center of the ovary (cloves).
The structure of the ovule before fertilization includes an achene (funiculus), a placenta, two integuments and a nucellus. The placenta is the place where the seed stalk attaches to the ovary. The inner part of the ovule - the nucellus - is covered with two integuments - integuments, external and internal. The integuments at the apex of the ovule do not close, forming a microscopic opening - the pollen duct, or micropyle. The part of the nucellus facing the micropyle is called the micropylar pole, and the opposite side is called the chalaza, or chalazal pole.
After a megaspore is formed from a nucellus cell, it grows in the ovule into the embryo sac, which occupies most of it, and the nucellus remains in the form of a small wall layer.
Fig.1 Gynoecium
Origin and evolution of the gynoecium
In the process of evolution of the gynoecium, the carpels gradually grow together and from the apocarpous gynoecium a coenocarpous gynoecium arises (from the Greek kainos - common). In the coenocarpous gynoecium, individual columns (stylodia) can remain free or grow together to form one common column (compound column). There are three types of coenocarpous gynoecium: syncarpous, paracarpous and lysicarpous.
Syncarpous (from the Greek syn - together) is a gynoecium consisting of a different number of closed carpels, fused together by lateral parts. It is bilocular and is characterized by the fact that the ovules are located along the sutures of closed carpels, i.e. at the angles formed by their abdominal parts (the so-called angular placentation). The syncarpous gynoecium usually derives from an apocarpous gynoecium with a cyclic (circular) arrangement of carpels, but in some cases it is also derived from a spiral gynoecium. A good example of a syncarpous gynoecium is the lily and tulip. At the first stages of the evolution of the syncarpous gynoecium, only the ovaries of the carpels grow together, and the styles (stylodia) remain free. But gradually the process of fusion also involves the columns, which eventually grow together into one complex column ending in a stigmatic head, which can be seen in heathers or in most monocots, including the lily.
Much more common than the syncarpous gynoecium is the paracarpous gynoecium (from the Greek para - near,). The paracarpous gynoecium is characterized by the openness of individual carpels while maintaining the connection of the marginal areas of neighboring carpels. In contrast to the syncarpous gynoecium, the paracarpous gynoecium is morphologically unilocular and is characterized by the so-called wall or parietal placentation (from the Latin parietalis - wall, Fig. 19). Well-known examples of paracarpous gynoecium are poppy, cucumber, and pumpkin. In many cases, the paracarpous gynoecium evolved from the syncarpous one. This was probably due to the fact that with the emergence of the syncarpous gynoecium, the need for the closure of each individual carpel in many at least cases becomes, in essence, unnecessary. Therefore, it is not surprising that the paracarpous gynoecium arose independently and in parallel in the most different lines of evolution of flowering plants and characterizes many families and entire orders. Structurally, the paracarpous gynoecium is more economical than the syncarpous one: with a relatively lower expenditure of building material, quite effective protection and supply of the ovules and the developing embryo is ensured. In addition, pollen tubes have wider access to the ovules than in the multilocular ovary of the syncarpous gynoecium.
In many other cases, the paracarpous gynoecium most likely evolved directly from the apocarpous one. This is most likely the origin of the paracarpous gynoecium of some African genera of the Annonaceae family, most of whose representatives are characterized by an apocarpous gynoecium, or the origin of the paracarpous gynoecium of the Canella family. The same, according to all data, is the origin of the paracarpous gynoecium of representatives of the families Savuraceae and Cactaceae, the entire order of Violaceae and a number of orders descended from it, as well as the family of Aquaticaceae among monocots.
In the paracarpous gynoecium, placentas usually grow, which often even branch. In many cases, these enlarged placentas collide in the center of the ovary, fuse with each other, and the cavity of the unilocular gynoecium is divided into chambers, which are false nests. Enlarged placentas are well expressed in some cucurbits, such as cucumber and watermelon.
In some lines of development of dicotyledons, for example, in purslanaceae and primroses, a lysicarpous gynoecium arose from the syncarpous gynoecium (from the Greek lysis - untying, liberation, dissolution). This is a single-locular gynoecium, in which the placentation is not parietal, as in the paracarpous type gynoecium, but the so-called free central, or columnar. The unilocularity of the lysicarpous gynoecium occurs in the process of evolution (or in ontogenesis) as a result of the disappearance of the septa (lateral walls) of the syncarpous ovary. In this case, the fused marginal parts do not collapse or diverge, and the ovules continue to sit on them. Thus, these remaining marginal parts of the carpels, together with the placentas, turn into a central column that rises in the cavity of the ovary.
In addition to these three main types of coenocarpous gynoecium, there is also a so-called pseudomonomeric gynoecium. This is a very reduced version of the syncarpous or paracarpous gynoecium, in which only one carpel is fertile and fully developed. Sterile carpels in a pseudomonomeric gynoecium are often so reduced that their presence can only be detected as a result of a special study of their anatomical structure and development. A typical pseudomonomeric gynoecium can be observed in elm, mulberry, nettle, hemp and related plants.
In many flowering plants, including primitive groups, the gynoecium is still free, not fused with the surrounding parts of the flower. But in many lines of evolution of flowering plants, completely independently and in parallel, a greater or lesser fusion of the gynoecium (usually coenocarpous) with the surrounding parts of the flower occurred, and as a result, the so-called inferior ovary arose. This process occurred gradually, and there are all intermediate forms between the upper and lower ovary. Therefore, in flower morphology, in addition to the terms “superior” and “lower” ovary, there is also the term “semi-inferior” ovary. The origin of the inferior ovary is one of the difficult questions in the evolutionary morphology of plants, and a large literature is devoted to it. To resolve this issue, the comparative anatomical study of the flower, especially its conducting system, was of great importance. The importance of the conducting system in solving various kinds of morphological issues is based on its relatively greater constancy (conservatism). Data on the anatomy of the conducting system of flowers with an inferior ovary lead to the conclusion that in the vast majority of cases, the inferior ovary occurred as a result of fusion of the bases of sepals, petals and staminate filaments with the ovary. The growth of a closely fitting floral tube to the ovary is quite natural and biologically justified. The different stages of this growth of the flower tube to the ovary can be traced, for example, in the heather family. However, as the same comparative anatomical studies have shown, the origin of the inferior ovary may be different. In some families, such as the Aizoonaceae, Cactaceae, Santalaceae and related families, as well as in some others, the ovary is not overgrown with a floral tube, but with a receptacle. The course of the vascular bundles clearly shows that the ovary seems to have sunk into the cup-shaped recess of the receptacle and fused with it. However, this second way of formation of the inferior ovary is observed only in a very small number of families.
Placentation and its types
Placentation is the method of arrangement of placentas and ovules (ovules) on carpels in the ovaries of flowering plants. Differences in placentation are important in understanding the evolution of flowering plants.
The location of placentas, as well as the placement of ovules, varies among different groups of flowering plants and is closely related to one or another type of gynoecium. There are two main types of placentation: sutural, or marginal, and laminal, or superficial.
In apocarpous gynoecium (including monomeric ones), sutural or marginal placentation is clearly expressed. In this case, the ovules are located along the edges of the carpels in two rows along the ventral sutures.
In coenocarpous gynoecium (syncarpnous, paracarpnous and lysicarpnous) there are different types of sutural placentation. This is due to the peculiarities of carpel fusion.
The syncarpous gynoecium is characterized by the fusion of conduplicate or involute carpels. They grow together in lateral parts, forming partitions that separate the formed ovary nests, the number of which corresponds to the number of fused carpels. The ventral sutures of the carpels bearing the placenta appear in the center of the ovary. This type of placentation is called central-marginal, or central-angular (lily, bellflower, borage).
In the paracarpous gynoecium, the carpels are fused not by their lateral surfaces, but by their edges, forming a single-locular ovary. Placentation in this type of gynoecium is wall-like, or pariental: wall-marginal if the placentas are located at the edges of the carpels and wall-dorsal if they are confined to the midribs of the carpels. It is characteristic of species of many families and orders (brassica, poppy, violet, willow, orchid, etc.).
The lysicarpous gynoecium also has a single-locular ovary, but the placentas in it are located in a column rising from the bottom of the ovary. This type of placentation is called columnar or axial (sometimes free central). Columnar placentation is characteristic of primroses and cloves.
Primitive angiosperms were characterized by laminal placentation, which is also found in some modern flowering plants (magnolia, lotus, susak, hornwort, water lily). This type of placentation is characterized by the attachment of ovules not along the ventral suture, but along the entire inner surface of the ovary or in certain places.
The place where the ovules are attached to the ovary is called the placenta. The placenta usually looks like a small swelling, outgrowth or protrusion formed by the tissues of the ovary.
Depending on the characteristics of the attachment of the ovules to the wall of the ovary, several types of placentation are distinguished (Fig. 2). The location of the ovules can be parietal, or pariental, when the ovules are located inside the ovary along its outer walls or in their protrusions. In the case when the ovules are located on the central column of the ovary, divided into nests according to the number of carpels, placentation is called axial or axial. With free central placentation, the ovules develop on a free central column, not connected by septa to the wall of the ovary. In some cases, the only ovule is located at the very base of a single-locular ovary. This type of placentation is called basal. There are other types of placentation, but they are relatively rare. The difference in placentation is of great importance in the taxonomy of flowering plants.

Fig. 2.: 1 - wall, or parietal, 2 - axial, or axial, 3 - free central, 4 - basal.
Ovary and its types
Ovary is a term of plant morphology; closed hollow container, the lower swollen part of the pistil of a bisexual or female flower. The ovary contains securely protected ovules. After fertilization, the ovary turns into a fruit, inside of which there are seeds developed from ovules.
The ovary acts as a moist chamber that protects the ovules from drying out, temperature fluctuations, and being eaten by insects.
The ovary and the stigma of the pistil, which serves to catch pollen, are connected by a column (if there are several pistils in a flower, their upper narrowed parts are called stylodes).
The ovary can be single-locular or multilocular (in the latter case it is divided by partitions into nests; sometimes the nests are separated by false partitions).
Based on the type of location in the flower, the ovaries are called:
Upper (free) ovary - attached by its base to the receptacle, without merging with any parts of the flower (in this case, the flower is called subpistillate or peripistotic).
The lower ovary is located under the receptacle, the remaining parts of the flower are attached at its apex (in this case the flower is called suprapistal).
Semi-inferior ovary - fuses with other parts of the flower, hypanthium or receptacle, but not at the very top, its apex remains free (in this case the flower is called semi-supapistal).

Fig.3. Types of ovaries
Structure and types of ovule
Ovule or ovule, a multicellular formation in seed plants from which the seed develops. The main parts of S. are the nucellus, the integument (or integuments), and the achene. The nucellus typically arises in the form of a tubercle from the cells of the placenta megasporophyll (carpel). The integument is formed in the form of an annular ridge at the base of the nucellus and overgrows the developing nucellus, leaving above its apex a narrow channel - the micropyle, or pollen passage, under which in most gymnosperms there is a pollen chamber. The seed stalk (funiculus) connects. with the placenta. the basal part from which the seed stalk, called a chalaza, extends.
In the ovary of the pistil there are small formations - ovules. Their number ranges from one (wheat, plum) to several million (in archidae). The functions of the ovule are megasporogenesis (creation of megaspores) and megagametogenesis (formation of the female gametophyte, fertilization process). The fertilized ovule develops into a seed. The placenta is the place where the ovule attaches to the carpel.
Parts of the ovule:
nucellus, ovule nucleus;
funiculus, the peduncle with which the ovule is attached to the placenta;
integuments, coverings of the ovule, forming a canal at the apex of the nucellus;
micropyle, pollen passage;
chalaza, the basal part of the ovule, where the nucellus and integuments merge;
hilum - the place where the ovule attaches to the peduncle.

Fig.4 Structure of the ovule
Types of ovule:
orthotropic - straight, funiculus and micropyle are located at opposite ends of the ovule axis (buckwheat, nut);
anatropic (reverse) - the nucellus is rotated 180 relative to the direct axis of the ovule, as a result of which the micropyle and funiculus are located nearby (Angiosperms)
hemitropic (semi-rotated) - the ovule is rotated by 90, as a result of which the micropyle and nucellus are located in relation to the funiculus at an angle of 90 (primroses, norica).
campylotropic (unilaterally curved) - the nucellus is curved unilaterally with a micropylar end, respectively, the micropyle and funiculus are located nearby (leguminous, malvaceae)
amphitropic (bilaterally curved) - the nucellus is curved bilaterally in the form of a horseshoe, with the micropyle and funiculus located nearby (mulberry, cistus).

Fig.5 Main types of ovules
6. Development of the ovule
Megasporogenesis occurs in the female reproductive sphere - in the gynoecium. Morphologically, the gynoecium is represented by a pistil (or pistils). The composition of the pistil includes: stigma, style and ovary. The ovary contains ovules (one or more). The internal contents of the ovule is a nucellus. The integument of the ovule is formed by a double or single integument. The nucellus of the ovule contains one archesporial cell (2n), capable of dividing by meiosis (in willows and some other plants the archesporium is multicellular). As a result of meiosis, four haploid megaspores (n) are formed from the archesporial cell (mother cell of the megaspores). Soon three of them die, and one increases in size and divides three times through mitosis. As a result, an eight-nucleated embryo sac (female gametophyte) is formed. Three nuclei, together with the adjacent cytoplasm, form antipodal cells, two nuclei - one central diploid nucleus; two nuclei - two synergid cells; one nucleus becomes the nucleus of the egg.
When a pollen tube approaches the ovule, it “feels” it in advance and prepares to meet it. Companion cells begin to secrete mucous substances. Meanwhile, the pollen tube grows, overcoming the resistance of the cells of the ovary walls. Finally, it reaches the micropyle. A “dramatic” process occurs: the pollen tube pierces (and in the process kills) one of the companion cells. Both sperm leave the pollen tube. The fate of the vegetative cell of a pollen grain is unenviable; it will soon die. It is very difficult to observe this process, but it is even more difficult to understand what happens during fertilization.
In August 1898, when double fertilization was not yet known, the Tenth Congress of Russian Naturalists and Doctors took place in Kyiv. Professor Sergei Gavrilovich Navashin made an important report on it: both sperm contained in the pollen grain are necessary for the normal development of seeds of two species from the Liliaceae family: lily (Lilium martagon) and hazel grouse (Fritillaria tenella). Why did Navashin choose these particular plants? Probably because their sperm and embryo sac are large, they are easy to examine under a microscope. Navashev was the first to state that the fertilization process involves two sperm merging with two (!) cells of the embryo sac. But let’s give the floor to the author of the discovery himself.
Each time a pollen tube was observed in contact with the embryo sac, both male sex nuclei were also observed in the contents of the embryo sac. Male nuclei lie close to each other at first...
The male nuclei then separate from each other, and one penetrates the egg, and the other is closely attached to one of the polar nuclei that has not yet merged at this time, namely to the sister nucleus of the egg...
While the male nucleus is more and more closely adjacent to the egg nucleus, the polar nucleus, copulating with another male nucleus, moves towards another polar nucleus, which it meets in the middle of the embryo sac...
Only after prophase of division do the nuclei merge..."
To make the explanation clearer, Sergei Gavrilovich made drawings that botanists in the capital really liked. They took them to show them at a meeting of the St. Petersburg Academy.
Why did scientists attach such great importance to this speech? It was believed that fertilization in plants occurs in the same way as in animals. One sperm and one egg should produce a zygote from which a new plant develops. It's not that botanists haven't seen two sperm moving in a pollen tube towards the embryo sac. It was believed that this was some kind of abnormality, a “typical” plant should have not two, but one sperm, and “typical” fertilization should be the same in both plants and animals. It turned out that these ideas were wrong. Fertilization of flowering plants is completely different from fertilization not only of animals, but also of other plants. The process of fertilization using two sperm is called double fertilization. Immediately after Navashin's discovery, scientists rushed to their drugs. It turned out that many had already seen double fertilization, but considered it an ugly process that supposedly did not produce normal seeds. Scientists began sending congratulations to Sergei Gavrilovich. And one scientist even donated his old preparations, which clearly showed double fertilization.
So, S.G. Navashin discovered an amazing phenomenon. It was clear why fertilization of the egg occurs: to create a zygote, and from it a new plant. But why fertilize the central cell of the embryo sac? It turned out that it is from this cell that the nutritional tissue of the seed, the endosperm, develops. Navashin suggested that endosperm cannot form without a male nucleus. How can we test this assumption? After all, it is impossible to remove one of the sperm from the pollen tract without disrupting the fertilization process.
Sergei Gavrilovich decided to study plants that do not have endosperm. What if fertilization of the central cell is disrupted? Such plants were found in the Orchid family (Orchidaceae). Orchid seeds are very small and cannot even germinate on their own (they germinate only with the help of basidiomycete fungi, forming mycorrhiza). Navashin saw two sperm cells in the orchid pollen tube. One of them fertilized the egg, and the second “tried” to fertilize the central cell with two polar nuclei. But in the central cell the nuclei did not merge with each other! The fertilization process was disrupted and, naturally, endosperm could not form. In further work, the scientist was able to show that in sunflower (Helianthus annuus) and some other plants, fertilization is double. After Navashin discovered double fertilization in a variety of plants, he concluded that double fertilization is characteristic of all flowering plants.
Let's take a look at the structure of the ovule after double fertilization. The cells of both of its integuments both had two sets of chromosomes and retained them. Both sets in these cells belong to the mother plant. Nucellus also carries two maternal sets of chromosomes. In the embryo sac, the synergids died, and the antipodes each had one set of chromosomes. These sets are also maternal ones. The zygote, formed by the fusion of an egg and a sperm, carries two sets of chromosomes: one from the paternal plant and the other from the maternal plant. The most interesting thing is that the central cell has three sets of chromosomes: one from the father plant, and two from the mother plant.
After double fertilization, several processes begin: the primary endosperm nucleus divides to form the endosperm, the zygote develops into the embryo, the integuments develop into the seed coat, and the ovary wall and associated structures form the fruit. At the early stages of development, the sequence of cell divisions in dicotyledonous and monocotyledonous embryos is similar; in both cases spherical bodies are formed. Later, a difference appears: the embryo of dicotyledons has two cotyledons, while that of monocotyledons has only one. In the seeds of some groups of angiosperms, the nucellus develops into storage tissue called the peristerm. Some seeds contain both endosperm and peristerm, as in beets (Belta). However, in many dicotyledons and some monocotyledons, all or most of the reserve tissue is absorbed by the developing embryo even before the seed enters a dormant state (in peas, beans, etc.). The embryos in such seeds usually develop fleshy cotyledons filled with nutrients. The main nutrients stored in seeds are carbohydrates, proteins, and lipids. The seeds of gymnosperms and angiosperms differ in the origin of these substances. In the former, they are produced by the female gametophyte, in the latter by the endosperm, which is neither gametophytic nor sporophytic tissue.
The development of the ovule into a seed is accompanied by the transformation of the ovary (and sometimes other parts of the plant) into a fruit. During this process, the wall of the ovary (pericarp, or pericarp) often thickens and differentiates into separate layers - the outer exocarp (extracarp), the middle mesocarp (intercarp) and the inner endocarp (intracarp) - usually more visible in fleshy than in dry fruits.
This is how the gametophyte develops, fertilization and seed formation occur in a “typical” flowering plant, but the plant world is very diverse and in many angiosperms development proceeds differently, but no less interesting and surprising. All these methods of development help each plant to maintain its strategy in the struggle for existence; The advantage of double fertilization is not limited to the fact that plants form nutritional reserves in the ovule only under the condition of fertilization (without “wasting” energy on the formation of nutritional reserves in the ovule, which will never be fertilized, as often happens in gymnosperms), these differences in development, which are a consequence of the adaptability of plants to their surrounding conditions, and create that amazing variety of forms of morphological and physiological structure that we observe in flowering plants.
7. Embryo sac and its development
The female gametophyte of flowering plants is usually called the embryo sac. The origin of this name goes back to those times when the morphological nature of the extremely peculiar female gametophyte of flowering plants was not yet clear. Currently, both terms are used as synonyms.

Fig.6. Embryo sac (female prothallus) of an angiosperm (flowering) plant and double fertilization: 1 - egg; 2 - helper cells; 3 - opened pollen tube. One of the sperm (4a) fuses with the egg; 5 - polar nuclei merging with the second sperm (46); 6 - three cells in the lower part of the embryo sac (antipodes).
In most flowering plants (probably at least 80%), the female gametophyte is initiated by one of four potential tetrad megaspores through three successive mitotic divisions. This type of formation of the female gametophyte is called monosporic. Its development occurs as follows. As a result of the first division of the nucleus of a functioning megaspore, two nuclei are formed, which diverge to the poles (micropylar and chalazal) of a strongly elongating megaspore, and a large vacuole is formed between them. Next, each of these two nuclei divides synchronously two more times and as a result, four nuclei are formed at each pole. This is the eight-core stage of female gametophyte development. From each of the two polar quadruples, one nucleus enters the central part of the embryo sac. These so-called polar nuclei come together in the central part and merge either immediately or only before fertilization, and sometimes do not merge at all. As a result of the fusion of polar nuclei, the nucleus of the central cell of the female gametophyte is formed, often called the secondary nucleus of the embryo sac.
Subsequently (during the last mitosis), cell partitions arise and the free-nuclear stage of development of the female gametophyte passes into the cellular stage. At the micropylar pole three cells of the egg apparatus are formed, at the opposite (chalazal) pole there are three cells called antipodes, and between these two groups of cells there is a central cell (containing two free polar nuclei or one secondary nucleus). The three cells of the egg apparatus differentiate into one egg and two cells, called synergids (from the Greek synergos - acting together) or auxiliary cells. This is the structure of the so-called normal monosporic female gametophyte.
The normal monosporic female gametophyte does not give the impression of a fully developed model in which economy has reached the same limit as in the case of the male gametophyte. The fact that the limit has not been reached here is proved by the following. In the process of evolution of this type of female gametophyte, the main type for flowering plants, embryo sacs of other types appeared, formed by two or even four megaspores. There was a reduction in mitotic divisions to two or one, and the distribution of nuclei also changed. Different combinations of these changes led to the emergence of several types of female gametophyte, which differ in both the number of nuclei (4, 8, 16), the number of cell groups and polar nuclei (1, 2, 4, 7-14), and the number of cells in groups (for example , the egg apparatus can consist of 1, 2, 3, 5 and 7 cells), as well as other characteristics.
The main feature of the monosporic type of formation of the female gametophyte is the sequential formation of cell partitions during meiotic divisions and the formation of separate megaspores. Megasporogenesis, which precedes the formation of bisporic (two-spore) and tetrasporic (four-spore) gametophytes, has a completely different character. Changes in megasporogenesis consist in the suppression of the formation of a cell septum during meiotic division, as a result of which, instead of tetrads of megaspores, non-cellular formations called coenocytes (from the Greek koinos - general and kytos - vessel, cell) appear, which further develop into female gametophytes. There are two possible options here.
In the first case, the formation of cell partitions is suppressed only during the second meiotic division. Therefore, both nuclei included in the viable cell of the dyad take part in the formation of the female gametophyte. A binuclear dyad is a coenocyte, the product of the fusion (or rather, non-separation) of two potential megaspores. Since each such coenocyte corresponds in origin to a pair of non-separated megaspores, it is considered as a two-spore coenocyte, and the female gametophyte developing from it is considered bisporic (two-spore). The bisporic female gametophyte is observed in onions, scilla, lily of the valley, amaryllis, chastuha and some other plants. The development of such a female gametophyte occurs in a more abbreviated manner and only two mitotic divisions are required to form the eight-nuclear stage.
Finally, in some plants, such as peperomia, elm, maynik, hazel grouse, tulip, lily, goose onion, plumbago, adoxa and others, the formation of cell partitions is suppressed during both meiotic divisions. The entire megaspore mother cell is transformed here into one four-spore coenocyte, or tetracenospore. A four-spore coenocyte corresponds to four undivided megaspores, which is why female gametophytes of this type are called tetrasporic (four-spore). To form the eight-nucleate stage of the female gametophyte, only one mitotic division of the nuclei of the four-spore coenocyte is required.
According to I.D. Romanov, in contrast to the monosporic female gametophyte, bi- and tetrasporic are called coenosporic.
The female gametophyte of flowering plants evolved from the more primitive archegonial female gametophyte of distant gymnosperm ancestors through the loss of the last stages of development leading to the formation of archegonia. And in coenosporic types, as we have seen, the initial stages also fall out. But while the evolution of the male gametophyte has reached its culmination stage and all possible initial and final stages have completely dropped out of its development, with the female gametophyte the evolutionary process continues to experiment. Hence a significant number of models of the female gametophyte and ways of its formation, which, however, there is no opportunity to dwell on here.
Literature
1. http://ru. wikipedia.org/wiki
2. I.I. Andreeva, L.S. Rodman "Botanica" / Moscow "Colossus" 2002
3. P. Raven, R. Evert, S. Eichhorn “Modern Botany” / Moscow “Mir” 1990
4. Mageshwari P., Embryology of Angiosperms, trans. from English, M., 1954; Takhtadzhyan A.L., Higher plants, vol. 1, M. - L., 1956;
5. Poddubnaya-Arnoldi V.A., General embryology of angiosperms, M., 1964;
6. Savchenko M.I., Morphology of the ovule of angiosperms, Leningrad, 1973.
The Chastukhova families can serve as good breeding material. The Chastuha family, which includes 13-14 genera and about 100 species, includes such widespread plants in Russia on the shores of reservoirs and swamps as common arrowhead (Sagittaria sagittifolia) with arrow-shaped leaves and clusters of rather large almost white flowers and plantain chastuha, or common ( Alisma...
... (dead water-conducting cells with lignified membranes), parenchyma living cells (carry out the transfer of photosynthesis products from the mesophyll and are adjacent to the phloem part of the bundle). 6.Anatomical structure of the root of monocots The root, like the stem, is a radially symmetrical organ characterized by apical growth, carried out by activity at its end of the meristem and involved in...


The shape of the shoot, or long leaves (in ferns). 2. Experimental work on the formation of educational activities in the process of studying the morphology of hanging plants in biology lessons 2.1 Methods for studying hanging plants in a school biology course Lesson objectives: to form the concept of a stem as the axial part of a modified shoot of hanging plants. Reveal the relationship between...
Quickly regenerates the outer shells of the aura, facilitating mutual understanding. 2. Deciduous Trees. Birch family (Betulaceae), genus Birch (Betula). There are 2 types of birch trees growing in the Murmansk region: silver birch (Betula pendula) and downy birch (Betula pubescens). For medicinal purposes, buds, leaves, bark, and birch tar are used. Birch leaf tea is the best remedy for...
Ovule or ovule, a multicellular formation in seed plants from which the seed develops. The main parts of S. are the nucellus, the integument (or integuments), and the achene. The nucellus typically arises in the form of a tubercle from the cells of the placenta megasporophyll (carpel). The integument is formed in the form of an annular ridge at the base of the nucellus and overgrows the developing nucellus, leaving above its apex a narrow channel - the micropyle, or pollen passage, under which in most gymnosperms there is a pollen chamber. The seed stalk (funiculus) connects. with the placenta. the basal part from which the seed stalk, called a chalaza, extends.
In the ovary of the pistil there are small formations - ovules. Their number ranges from one (wheat, plum) to several million (in archidae). The functions of the ovule are megasporogenesis (creation of megaspores) and megagametogenesis (formation of the female gametophyte, fertilization process). The fertilized ovule develops into a seed. The placenta is the place where the ovule attaches to the carpel.
Parts of the ovule:
● nucellus, ovule nucleus;
● funiculus, the peduncle with which the ovule is attached to the placenta;
● integuments, coverings of the ovule, forming a canal at the apex of the nucellus;
● micropyle, pollen passage;
● chalaza, the basal part of the ovule, where the nucellus and integuments merge;
● hilum - the place where the ovule attaches to the peduncle.
Rice. Structure of the ovule
Types of ovule:
orthotropic - straight, funiculus and micropyle are located at opposite ends of the ovule axis (buckwheat, nut);
anatropic (reverse) - the nucellus is rotated 180 relative to the direct axis of the ovule, as a result of which the micropyle and funiculus are located nearby (Angiosperms)
hemitropic (semi-rotated) - the ovule is rotated by 90, as a result of which the micropyle and nucellus are located in relation to the funiculus at an angle of 90 (primroses, norica).
campylotropic (unilaterally curved) - the nucellus is curved unilaterally with a micropylar end, respectively, the micropyle and funiculus are located nearby (leguminous, malvaceae)
amphitropic (bilaterally curved) - the nucellus is curved bilaterally in the form of a horseshoe, with the micropyle and funiculus located nearby (mulberry, cistus).

Rice. Main types of ovules
Development of the ovuleMegasporogenesis occurs in the female reproductive sphere - in the gynoecium. Morphologically, the gynoecium is represented by a pistil (or pistils). The composition of the pistil includes: stigma, style and ovary. The ovary contains ovules (one or more). The internal contents of the ovule is a nucellus. The integument of the ovule is formed by a double or single integument. The nucellus of the ovule contains one archesporial cell (2n), capable of dividing by meiosis (in willows and some other plants the archesporium is multicellular). As a result of meiosis, four haploid megaspores (n) are formed from the archesporial cell (mother cell of the megaspores). Soon three of them die, and one increases in size and divides three times through mitosis. As a result, an eight-nucleated embryo sac (female gametophyte) is formed. Three nuclei, together with the adjacent cytoplasm, form antipodal cells, two nuclei - one central diploid nucleus; two nuclei - two synergid cells; one nucleus becomes the nucleus of the egg.
When a pollen tube approaches the ovule, it “feels” it in advance and prepares to meet it. Companion cells begin to secrete mucous substances. Meanwhile, the pollen tube grows, overcoming the resistance of the cells of the ovary walls. Finally, it reaches the micropyle. A “dramatic” process occurs: the pollen tube pierces (and in the process kills) one of the companion cells. Both sperm leave the pollen tube. The fate of the vegetative cell of a pollen grain is unenviable; it will soon die. It is very difficult to observe this process, but it is even more difficult to understand what happens during fertilization.
In August 1898, when double fertilization was not yet known, the Tenth Congress of Russian Naturalists and Doctors took place in Kyiv. Professor Sergei Gavrilovich Navashin made an important report on it: both sperm contained in the pollen grain are necessary for the normal development of seeds of two species from the Liliaceae family: lily (Lilium martagon) and hazel grouse (Fritillaria tenella). Why did Navashin choose these particular plants? Probably because their sperm and embryo sac are large, they are easy to examine under a microscope. Navashev was the first to state that the fertilization process involves two sperm merging with two (!) cells of the embryo sac. But let’s give the floor to the author of the discovery himself.
Each time a pollen tube was observed in contact with the embryo sac, both male sex nuclei were also observed in the contents of the embryo sac. The male nuclei initially lie close to each other.
The male nuclei then separate from each other, and one penetrates the egg, and the other is closely attached to one of the polar nuclei that has not yet merged at this time, namely the sister nucleus of the egg.
While the male nucleus is more and more closely attached to the egg nucleus, the polar nucleus, copulating with another male nucleus, moves towards another polar nucleus, which it meets in the middle of the embryo sac.
Only after prophase of division do the nuclei merge..."
To make the explanation clearer, Sergei Gavrilovich made drawings that botanists in the capital really liked. They took them to show them at a meeting of the St. Petersburg Academy.
Why did scientists attach such great importance to this speech? It was believed that fertilization in plants occurs in the same way as in animals. One sperm and one egg should produce a zygote from which a new plant develops. It's not that botanists haven't seen two sperm moving in a pollen tube towards the embryo sac. It was believed that this was some kind of abnormality, a “typical” plant should have not two, but one sperm, and “typical” fertilization should be the same in both plants and animals. It turned out that these ideas were wrong. Fertilization of flowering plants is completely different from fertilization not only of animals, but also of other plants. The process of fertilization using two sperm is called double fertilization. Immediately after Navashin's discovery, scientists rushed to their drugs. It turned out that many had already seen double fertilization, but considered it an ugly process that supposedly did not produce normal seeds. Scientists began sending congratulations to Sergei Gavrilovich. And one scientist even donated his old preparations, which clearly showed double fertilization.
So, S.G. Navashin discovered an amazing phenomenon. It was clear why fertilization of the egg occurs: to create a zygote, and from it a new plant. But why fertilize the central cell of the embryo sac? It turned out that it is from this cell that the nutritional tissue of the seed, the endosperm, develops. Navashin suggested that endosperm cannot form without a male nucleus. How can we test this assumption? After all, it is impossible to remove one of the sperm from the pollen tract without disrupting the fertilization process.
Sergei Gavrilovich decided to study plants that do not have endosperm. What if fertilization of the central cell is disrupted? Such plants were found in the Orchid family (Orchidaceae). Orchid seeds are very small and cannot even germinate on their own (they germinate only with the help of basidiomycete fungi, forming mycorrhiza). Navashin saw two sperm cells in the orchid pollen tube. One of them fertilized the egg, and the second “tried” to fertilize the central cell with two polar nuclei. But in the central cell the nuclei did not merge with each other! The fertilization process was disrupted and, naturally, endosperm could not form. In further work, the scientist was able to show that in sunflower (Helianthus annuus) and some other plants, fertilization is double. After Navashin discovered double fertilization in a variety of plants, he concluded that double fertilization is characteristic of all flowering plants.
Let's take a look at the structure of the ovule after double fertilization. The cells of both of its integuments both had two sets of chromosomes and retained them. Both sets in these cells belong to the mother plant. Nucellus also carries two maternal sets of chromosomes. In the embryo sac, the synergids died, and the antipodes each had one set of chromosomes. These sets are also maternal ones. The zygote, formed by the fusion of an egg and a sperm, carries two sets of chromosomes: one from the paternal plant and the other from the maternal plant. The most interesting thing is that the central cell has three sets of chromosomes: one from the father plant, and two from the mother plant.
After double fertilization, several processes begin: the primary endosperm nucleus divides to form the endosperm, the zygote develops into the embryo, the integuments develop into the seed coat, and the ovary wall and associated structures form the fruit. At the early stages of development, the sequence of cell divisions in dicotyledonous and monocotyledonous embryos is similar; in both cases spherical bodies are formed. Later, a difference appears: the embryo of dicotyledons has two cotyledons, while that of monocotyledons has only one. In the seeds of some groups of angiosperms, the nucellus develops into storage tissue called the peristerm. Some seeds contain both endosperm and peristerm, as in beets (Belta). However, in many dicotyledons and some monocotyledons, all or most of the reserve tissue is absorbed by the developing embryo even before the seed enters a dormant state (in peas, beans, etc.). The embryos in such seeds usually develop fleshy cotyledons filled with nutrients. The main nutrients stored in seeds are carbohydrates, proteins, and lipids. The seeds of gymnosperms and angiosperms differ in the origin of these substances. In the former, they are produced by the female gametophyte, in the latter by the endosperm, which is neither gametophytic nor sporophytic tissue.
The development of the ovule into a seed is accompanied by the transformation of the ovary (and sometimes other parts of the plant) into a fruit. During this process, the wall of the ovary (pericarp, or pericarp) often thickens and differentiates into separate layers - the outer exocarp (extracarp), the middle mesocarp (intercarp) and the inner endocarp (intracarp) - usually more visible in fleshy than in dry fruits.
This is how the gametophyte develops, fertilization and seed formation occur in a “typical” flowering plant, but the plant world is very diverse and in many angiosperms development proceeds differently, but no less interesting and surprising. All these methods of development help each plant to maintain its strategy in the struggle for existence; The advantage of double fertilization is not limited to the fact that plants form nutritional reserves in the ovule only under the condition of fertilization (without “wasting” energy on the formation of nutritional reserves in the ovule, which will never be fertilized, as often happens in gymnosperms), these differences in development, which are a consequence of the adaptability of plants to their surrounding conditions, and create that amazing variety of forms of morphological and physiological structure that we observe in flowering plants.
ovule ovule
ovule, ovule (ovulum), multicellular formation of seed plants, from which the seed develops. Mn. Embryologists consider S. a structure homologous to the megasporangium of spore plants. C consists of a nucellus containing a megasporocyte, two or one integuments (covers), which, when closed, form a narrow channel - micropyle, through which the pollen tube penetrates to the embryo sac, as well as a funiculus (ovules), which attaches S. to the placenta. The part opposite the micropyle is called S. chalaza. S. of flowering plants is formed in the ovary. S. gymnosperms are naked, sitting on megasporophyll. In the chalazal part of S., as a result of cell differentiation, a hypostasis is formed. For plural Flowering plants in S. are characterized by obturators - sections of tissue that grow in the form of papillae towards the micropyle and facilitate the penetration of the pollen tube into the embryo sac, its growth and nutrition. S. is formed on the placenta in the form of a meristematic tubercle. cells from outside epidermis layer; near its apex one or two archesporial cells (archesporia) appear from the subepidermal layer, and at the base integuments appear in the form of one or two annular ridges. The megasporocyte (mother cell of the megaspores) gives rise to megaspores, of which the lower (chalazal), less often the upper (micropylar) give rise to females. gametophyte (embryo sac in flowering plants or primary endosperm in gymnosperms). After fertilization, with the beginning of the development of the embryo, S. turns into a developing seed. There are 5 main ones. types of S. depending on the location of the micropyle, funiculus and longitudinal axis of the nucellus: orthotropic, or straight (in buckwheat, pepper, aroid); anatropic, or reversed (the most common type, perhaps the original); hemitropic, or semi-rotated (in some noricaceae, in primroses); campylotropic, or one-sided curved (in many species of cloves, legumes, etc.); amphitropic, or bilaterally curved (in some species of the same orders as the previous type). S. with a powerful nucellus, thick, sometimes lignifying integuments (the so-called crassinucellate) is considered primitive (prevails in gymnosperms), with a weakly expressed nucellus (tenuinucellate) and with one integument - more progressive, arising from the first by gradual reduction (prevails in flowering). The number of S. in the ovary of flowering plants ranges from one (in cereals) to 1 million (in orchids).
.(Source: “Biological Encyclopedic Dictionary.” Editor-in-chief M. S. Gilyarov; Editorial Board: A. A. Babaev, G. G. Vinberg, G. A. Zavarzin and others - 2nd ed., corrected - M.: Sov. Encyclopedia, 1986.)
ovule(ovule), a multicellular formation in the ovary of seed plants, from which the seed develops after fertilization. Usually consists of outer and inner integuments (instruments). They do not close, leaving a narrow opening - the pollen duct (micropyle). The integument covers a multicellular closed layer - the nucellus, which encloses the embryo sac. It, in turn, consists of an egg apparatus - three cells concentrated at the end closest to the pollen tube. One of them, with a larger nucleus, is the egg (female gamete), the other two are auxiliary cells, or syneriguda. At the end opposite the pollen tube, 3 antipodal cells develop. In the middle of the embryo sac there is a central cell. The egg cell and the central cell are involved in fertilization.
.(Source: “Biology. Modern illustrated encyclopedia.” Chief editor A. P. Gorkin; M.: Rosman, 2006.)
Synonyms:
See what "ovule" is in other dictionaries:
The ovules of cycads differ in size (from 5-6 cm in length in some cycad species to 5-7 mm in dwarf zamia) and in shape. But at the same time, they are quite similar in the main features of development and internal structure. Openly seated... ... Biological encyclopedia
Ovule Dictionary of Russian synonyms. ovule noun, number of synonyms: 2 ovule (2) ... Synonym dictionary
Same as ovule... Big Encyclopedic Dictionary
Weaving; m. Botan. Seed embryo; ovule. * * * ovule is the same as ovule. * * * ovule ovule, the same as ovule (see ovule) ... encyclopedic Dictionary
The location of the ovules in the flower of the stinking hellebore (Helleborus foetidus) Ovule, or ovule (lat. ... Wikipedia
Ovule ovule, ovule. The multicellular organ of seed plants, from which the seed develops, is formed in the ovary in angiosperms; S. consists of nucellus
ovule- Synonyms: ovule of megasporangium of seed plants, surrounded by one or two integuments - integuments. It is formed on megasporophylls or in female strobili (cones) of gymnosperms, and in angiosperms - inside the ovary of the pistil of a flower (see ... Anatomy and morphology of plants
Same as ovule... Natural science. encyclopedic Dictionary
ovule- seed atok, weave... Russian spelling dictionary
ovule- ovule (ovulum), a multicellular formation in the reproductive organs of seminal cells, from which the seed develops during development (usually after fertilization). In angiosperms, S. is formed hidden inside the ovary; in gymnosperms it is located... ... Agricultural Encyclopedic Dictionary
- In contact with 0
- Google+ 0
- OK 0
- Facebook 0