Gastrulation in the lancelet represents a convenient model for studying the general patterns of this process in the chordate phylum. This process, as was shown earlier in the lancelet, occurs according to the type of intussusception. At the early stage of the gastrula, two germ layers are found: the ectodermal layer, formed from the roof of the blastula, and the endodermal layer, from the bottom of the blastula. The gastrocoel communicates with the external environment via a blastopore.
The next stage of embryo development begins - the formation of the third germ layer - mesoderm. The cells of the dorsal lip of the blastopore, rapidly multiplying, are immersed in the endoderm and on its dorsal side form a notochordal plate, the rudiment of the future notochord. Small cells of the ventral and lateral lips of the blastopore also grow into the endodermal layer and are located in its composition on the sides of the notochordal plate.
The two-layer embryo takes on an elongated shape, containing the material of three germ layers. Mesoderm material occupies the dorsal part of the inner layer. The neural plate emerges from the primary ectoderm located above the notochordal plate. The latter sinks and forms a neural groove along the length of the embryo. The neural groove, closing upward, forms the neural tube.
The notochordal plate emerges from the primary inner layer, closes downwards and forms a dense cellular cord - the notochord.
At the same time, the small cell material, located in the primary inner layer on the sides of the notochord, protrudes in the dorsal direction in the form of two pockets. Initially, these pockets open into the gastrocoel and then separate into two closed folds located along the gastrula.
This is how the third germ layer is formed - the mesoderm. After the separation of the notochord and mesoderm, the material of the primary inner layer closes, forming the endoderm - the inner germ layer (Fig. 5).
GASTRULATION in birds
Gastrulation in birds represents a fairly close model to the processes occurring in higher chordates, including humans.
In birds, a blastodisc is formed as a result of discoidal (meroblastic) fragmentation. It consists of irregularly shaped cells, tightly adjacent to each other and located on a huge mass of uncrushed yolk. Under the influence of enzymes from the blastodisc cells, part of the yolk liquefies and a cavity filled with liquid is formed - the blastocoel. The roof of this blastula is represented by a blastodisc, the bottom by an uncrushed mass of yolk. Blastocoel - subembryonic cavity.
Then comes gastrulation, consisting of two phases. The first phase begins before the egg is laid and consists of the separation of the endoderm by delamination of the blastodisc cells. As a result, a two-layer embryo appears. The upper layer of discoblastula cells takes on a prismatic shape and is arranged in one regular row. The lower layer of discoblastula cells retains a round or irregular shape and lies randomly on the yolk. A cavity appears between the cells of the upper and lower layers - the gastrocoel.
The second phase of gastrulation begins in the second half of the first day of incubation. The blastodisc grows, in the center of it a germinal shield is released, from which the body of the embryo subsequently develops. The part of the discoblastula surrounding the embryonic shield represents extra-embryonic material, in which a light and dark field is distinguished. The light field is located around the embryonic shield and consists of cells separated from the yolk by the subembryonic cavity due to the partial use of the yolk by the embryo. The dark field occupies the periphery of the discoblastula and consists of cells tightly adjacent to the yolk and growing along its surface. At this point, the division of the cellular material into the embryonic (embryonic shield) and extraembryonic (light and dark field) part is clearly expressed.
During 24 hours of incubation or incubation on the germinal shield, as a result of cell proliferation, they are observed to move from front to back along the periphery of the germinal shield. Both streams of cells meet centrally at the posterior end of the germinal shield, merge and move anteriorly in the middle of the germinal shield. As a result, a thickened strand of cells is formed, called the primitive streak.
At the anterior end of the primary stripe, a thickening is formed - the primary nodule. Subsequently, part of the blastomeres located anterior to the primary node moves to the latter, tucks under the ectoderm and forms a cephalic or notochordal outgrowth growing anteriorly between the ectoderm and endoderm - the rudiment of the notochord.
The blastomeres of the posterior half of the embryonic shield are shifted to the primitive streak and through this area are immersed under the ectoderm, located in the cavity between the ectoderm and endoderm. This material forms the rudiment of mesoderm - the third germ layer. Cells of the notochordal process multiply and... move, occupying a central position between the ectoderm

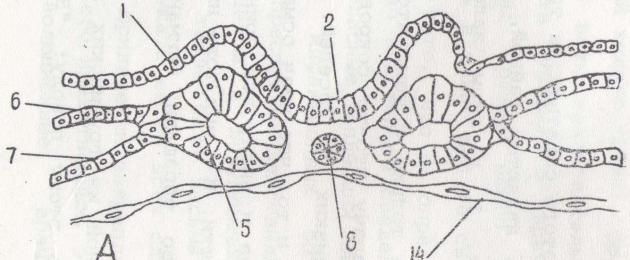
Fig.%6. Development of a complex of axial organs in birds 1-ectoderm; 2 - neural groove; 3 - neural tube; 4 - amnion; 5 - somites; 6 - parietal leaf of splachnotome; 7 - visceral leaf of the splanchnotome; 8 - chord; 9 - dermatome; 10 - myotome; 11 - sclerotome; 12 - nephrotoma; 13 - aorta; 14 - endoderm; 15 - amniotic fold and endoderm in the anterior half of the embryonic shield. Following this, mesoderm cells also move to the anterior half of the embryonic shield, located between the ectoderm and endoderm on the sides of the chordal outgrowth. This is parachordal mesoderm. It can be divided into dorsal and ventral.
As the cells of the primitive streak move into the anlage of the notochord and mesoderm, the material of the ectoderm and the neural plate included in it remains on the surface. Thus, the primary streak and the head node correspond in importance to the blastopore of the lancelet, since in this place the movement of germinal material takes place, leading to the formation of 3-germ layers. The rudiment of the intestinal endoderm is located most deeply. It is directly adjacent to the surface of the yolk and is located in the inner layer of the discoblastula.
By the end of gastrulation, the body of the embryo contains all the rudiments, from which the complex of axial organs typical of chordates arises (Fig. 6.).
The result of active cell division, growth and directed movements (migration) of cell flows with the formation of a multilayer embryo, or gastrula (the appearance of layer-by-layer germ layers, separated from each other by a distinct gap: outer - ectoderm, middle - mesoderm, inner - endoderm). The movement of cells occurs in a strictly defined area of the embryo - in the gray crescent area. The latter was described by V. Roux in 1888. In a fertilized amphibian egg, the gray sickle is revealed as a colored area on the side opposite to the penetration of the sperm. Factors necessary for gastrulation are believed to be localized in this location.
In different representatives of vertebrates it occurs in several main ways: by invagination (invagination), immigration (movement of some cells inside the embryo), epiboly (fouling), delamination (splitting). The methods of gastrulation depend on the type of egg. With any method of gastrulation, the leading forces are the uneven proliferation of cells in different parts of the embryo, the level of metabolic processes in cells located in different parts of the embryo, the activity of amoeboid cell movements, as well as inductive factors (proteins, nucleoproteins, steroids, etc.). As a result of gastrulation, the main rudiments of organs and tissues are separated.
Next period embryogenesis is histo- and organogenesis - differentiation of various tissues and organs of the body from the material of germ layers and embryonic rudiments.
As a result of gastrulation, a multilayered embryo. Despite the different methods of gastrulation, after the material of the germ layers is released, along the axis of the embryo there is the material of the notochord, which underlies the neural plate; to the left and right of the notochord is the material of the mesoderm. All this characterizes the axial complex of rudiments. Subsequently, the formation of organ rudiments occurs, which are spatially localized groups of stem cells - sources of tissue development. The patterns of differentiation of the cellular material of the primordia can be traced in the embryogenesis of the most studied animals.
Lancelet. Development of the lancelet.
A classic object of embryological research lanceentula, studied in detail by A.O. Kovalevsky. The lancelet is a representative of the class of chordates of the skullless subtype, up to 8 cm in size and lives on the sandy bottom in warm seas. It got its name because of its shape, reminiscent of a lancet (a surgical instrument with a double-edged blade, a modern scalpel).
Egg lancelet is oligo- and isolecithal, 110 µm in size, the nucleus is located closer to the animal pole. Fertilization is external. The fragmentation of the zygote is complete, almost uniform, synchronous and ends with the formation of a blastula. As a result of the alternation of meridian and latitudinal cleavage furrows, a single-layer blastula is formed with a cavity filled with fluid - the blastocoel. The blastula retains polarity, its bottom represents the vegetative part, and the roof represents the animal part; between them there is a marginal zone.
At gastrulation the vegetative part of the blastula invaginates into the animal part. The invagination gradually deepens and, finally, a double-walled cup is formed with a wide gaping hole leading into the newly formed embryonic cavity. This method of gastrulation is called intussusception. This is how the blastula turns into a gastrula. In it, the material of the embryo turns out to be differentiated into an outer layer - ectoderm, and an inner layer - endoderm. The cavity of the cup is called the gastrocoel, or the cavity of the primary intestine, which communicates with the external environment through a blastopore, which corresponds to the anus. The blastopore has a dorsal, ventral and two lateral lips. As a result of invagination, the center of gravity of the embryo shifts, and the embryo turns upward with its blastopore. Gradually, the edges of the blastopore close and the embryo elongates. The topography of cells within the blastopore lips determines the development of different parts of the embryo. During gastrulation, the notochord and mesoderm are separated from the inner layer of the gastrula, which are located between the ecto- and endoderm. Gastrulation ends with the formation of the axial complex of rudiments and then with the separation of organ rudiments. The notochord induces the development of a neural tube from dorsal ectoderm material. This part of the ectoderm thickens, forming a neural plate (neuroectoderm), which bends along the midline and turns into a groove.

The edges of the groove gradually close into the neural tube. Remaining part ectoderm- cutaneous, grows together over the neural tube. However, at the very anterior and posterior ends of the embryo, the neural tube communicates for some time with the external environment through two openings - neuropores. Subsequently, the mesoderm is divided into dorsal segments - somites, the number of which increases from 15 pairs to 60-65 pairs in an adult lancelet. Part of the laterally located mesoderm is not segmented and splits into outer (parietal) and inner (visceral) layers of the splanchnotome. These leaves grow between the ecto- and endoderm and, having reached the middle on the ventral side of the embryo under the intestinal tube, grow together, forming a single secondary cavity - the coelom. At the anterior end of the embryo, a depression (oral bay) appears, growing towards the anterior section of the intestinal tube. When the ectoderm of the oral bay comes into contact with the blind end of the intestinal tube, cell apoptosis occurs and communication between the intestine and the external environment occurs. A similar process occurs at the posterior end of the embryo. On the sides of the head section of the embryo, contact between the skin ectoderm and the intestinal endoderm also occurs. A breakthrough occurs at the point of this contact. This is how the cavity of the foregut communicates with the external environment (the gill apparatus is formed). After this, the embryo emerges from the egg shell into the external environment in the form of a larva.
Labeling methods for studying migration processes blastomeres made it possible to identify certain areas of the embryo in the early stages of development (zygotes - blastula), which later develop into germ layers and embryonic rudiments of organs and tissues. These areas were called presumptive areas, or rudiments.
The development of the lancelet was first studied by A. O. Kovalevsky. This question is of great interest, since analysis of the stages of development of the most primitive of modern chordate animals provides some basis for judging the early stages of chordate phylogeny. In addition, lancelet development provides a simplified schematic picture of embryonic development in other chordates. Figure 4 and Figure 5 depict the successive stages of embryonic development of the lancelet up to the formation of the larva. The fragmentation of the fertilized egg is complete and almost uniform: when a blastula is formed, it is clear that on its lower side, corresponding to the vegetative ("plant") part of the egg, the cells are larger than on the upper. Because of this, the inner layer of the next gastrula stage is represented by larger cells. Crushing occurs very quickly. In the ectoderm of the upper side of the embryo, a medullary plate is separated, the edges of which curl up and then close. The neural tube arising in this way maintains communication with the external environment at the anterior end (through the neuropore), and at the posterior end (through the neurointestinal canal) with the cavity of the gastrula, i.e. with the primary gut. Subsequently, the neurointestinal canal disappears completely, and the olfactory fossa remains in place of the neuropore.
From the lateral plate the peritoneum, mesenteries (in which the main blood vessels arise in the form of longitudinal canals), and intestinal muscles develop. Nephridial tubules develop as finger-like protrusions of the walls of the secondary body cavity. The gonads develop as protrusions of that part of the walls of the body cavity that corresponds to the place of separation of the somite and the lateral plate of the gonotome. The mouth is formed by protrusion of the primary intestine at the end opposite the gastropore (primary mouth) and counter-invagination of the ectoderm. A breakthrough occurs at the meeting point of these formations. The formation of the mouth and gill slits occurs asymmetrically. The oral opening is formed on the lower left side of the embryo. The left gill slits (there are 14 of them) initially appear on the ventral side and then move to the right side of the embryo. Then another row of slits appears here (there are 8 of them), located above the previously mentioned 14 slits. Subsequently, the lower row of slits shifts to the abdominal side and only after that to the left side of the body. Their number is reduced from 14 to 8. The number of gill slits on both sides then increases sharply. Subsequently, the mouth moves to the ventral side. The atrial cavity appears initially in the form of a groove on the lower surface of the body. The metapleural folds that form this groove grow towards each other and, closing, form a cavity that opens outward only in its posterior part, where the mentioned folds do not grow together. In general, the larval development of the lancelet lasts about three months.
Vertebrates evolved from skullless animals. The modern representative of the skullless subtype is the lancelet (Fig. 34). In the development of the lancelet we see the simplest scheme of embryonic development of chordates, which became significantly more complex during the process of evolution in vertebrates and especially in humans.
The lancelet is a marine animal. The female and male release reproductive cells (eggs and sperm) directly into the water, where fertilization and development of the embryo occurs.
Following fertilization, the zygote enters a period of cleavage (Fig. 35); the number of blastomeres increases rapidly (2, 4, 8, 16, etc.).

During the process of division, blastomeres gradually move away from the center of the embryo to the periphery, forming an ever-increasing cavity in the center.
In this regard, by the end of the crushing period, a typical blastula appears, the wall of which is formed by one layer of cells (blastoderm), and its cavity (blastocoel) is filled with fluid. The next period (gastrulation) is associated with intussusception, i.e., invagination of one (vegetative) half of the blastula into the other (animal) * (Fig. 35). As a result, a gastrula ** is formed, which has an internal germ layer (primary endoderm), a gastrocoel (primary gut cavity) and a blastopore (primary mouth).
* (In a lancelet egg, one half contains more yolk than the other. As a result of studying the development of embryos, it was established that the part of the egg, supplied with a large amount of yolk, when crushed, forms that half of the blastoderm, which invaginates during the gastrulation period and forms the internal germ layer - the endoderm. It is known that the digestive and other systems of the so-called plant (vegetative) life are formed from the endoderm. Therefore, both the part of the egg containing a larger amount of yolk and the part of the blastula formed from it as a result of crushing are called vegetative parts. From the opposite part of the egg and the corresponding part of the blastula, the ectoderm and then the organs of the animal part develop, in particular the nervous system, etc. Therefore, these parts of the egg and blastula are called animal.)
** (Greek, gaster - stomach. Hence the name “gastrula” to emphasize that the embryo at this stage is equipped with the rudiment of the digestive system in the form of the primary intestine.)
During the period of invagination, the blastocoel (primary body cavity) remains for some time in the form of a narrow gap between the outer and inner germ layers, and then disappears. After formation, the gastrula begins to increase in length; at the same time, concentric closure of the edges of the blastopore (primary mouth) * occurs.
* (The blastopore (primary mouth), connecting the primary gut with the external environment, in some animals at subsequent stages of development remains as an oral opening (protostomes), while in others it becomes an anal opening (deuterostomes). In the latter case, the oral opening is formed at the opposite end. Protostomes include worms, mollusks and arthropods, deuterostomes include echinoderms and chordates, in particular the lancelet and vertebrates, including humans.)
The end of gastrulation coincides with the beginning of the period of separation of the main rudiments of organs and tissues (Fig. 36). At this time, the thickened dorsal portion of the primary ectoderm turns into the neural plate, from which the neural tube arises, passing through the neural groove stage (Fig. 36, 37). The neural tube is the rudiment of the nervous system.


The other part of the outer germ layer, during further development, closes over the neural tube and is the rudiment of the skin epithelium (epidermis).
The inner germ layer undergoes a number of changes in cellular composition, then the following formations separate from it: in the area of the middle part of its roof - a notochordal plate, from which the notochord rudiment is formed; in the area of the lateral parts of the roof of the primary intestine there are pocket-shaped protrusions, which are then separated from the wall of the primary intestine. The cellular material of the detached pocket-like protrusions of the primary intestine fills the primary body cavity (located between the ectoderm and endoderm) and represents the rudiment of the middle germ layer (mesoderm). In the center of the isolated area there is a space detached from the cavity of the primary intestine, which is the secondary body cavity. The remaining part of the primary endoderm (bottom of the primary intestine) forms the intestinal tube, which is the rudiment of the secondary (definitive) intestine and from which the intestinal epithelium subsequently arises.
Thus, by the end of gastrulation, during the period of separation of the rudiments of organs and tissues, the neural tube is located on the dorsal side of the embryo in the middle position, and under it the notochord and the intestinal tube are successively located. Bilateral symmetry is finally revealed. Mesodermal pockets appear. The rudiments of mesoderm with cavities inside them grow on the right and left into the gap between the skin ectoderm and the intestinal tube and connect under the latter. At the same time, the mesoderm, extended along the body between the cutaneous ectoderm, on the one hand, and the notochord and intestinal tube, on the other hand, is divided into a number of separate sections (segments) located along the length of the body near each other. The left and right mesodermal pockets are subject to segmentation. At the same time, each mesodermal sac is divided throughout its entire length into a dorsal section (somite) and a ventral section (splanchnotome). The somites lose their cavity, become dense and serve as the starting material for the trunk muscles. Splanchnotomes retain a cavity. They remain separated from each other for some time (as a result of segmentation), and then the isolated cavities contained in separate splanchnotomes merge, so that a coelomic cavity (secondary body cavity, coelom) is formed for all segments of the body. The material of the walls of splanchnotomes is the original material of the epithelial lining of the secondary body cavity *.
* (The secondary body cavity (coelom) arises in the thickness of the middle germ layer. Various cavities are formed from it during embryonic development. In humans, these are, in particular, the peritoneal and pleural cavities and the pericardial cavity. The primary body cavity (blastocoel) disappears during gastrulation and the formation of mesoderm.)
The development of the lancelet larva ends with the appearance of the oral and anal openings, gill slits, the formation of organs, etc.
Features of the embryonic development of anamnia are studied using the example of lancelet, fish and amphibians.
The eggs of the lancelet are primary isolecithal , fertilization takes place in water, i.e. external After fertilization, a zygote is formed, which undergoes complete and uniform fragmentation - development holoblastic . The zygote is divided first by two successive mitoses in mutually perpendicular meridional planes into four, then by the equatorial furrow into eight blastomeres, etc. The cleavage planes alternate, and after the seventh division a blastula of the type appears coeloblastula .
Blastomeres, forming blastoderm vary in size and quality, because... there is a distribution of different quality material in the cytoplasm of the zygote, which undergoes internal differentiation. The resulting coeloblastula consists of large-cell yolk blastomeres forming the bottom (future intestinal endoderm), medium-sized blastomeres located dorsally above them - the material of the dorsal falx (future notochord) and small blastomeres surrounding the bottom of the blastula - the material of the central falx (future mesoderm). All this is surrounded by ectoderm.
Using the intravital staining method, it was found that all of the listed areas of the blastula move by tucking through the lips of the blastopore, are located around the gastrocoel and create the basis for the organotypic period of lancelet development - the period of differentiation of tissues and organs.
The blastula has a cavity - blastocoel . The blastocoel is filled with liquid - a waste product of blastoderm cells.
By intussusception , i.e. retraction of the vegetative hemisphere into the animal one, the blastula is transformed into gastrulu , the wall of which becomes two-layer and consists of ectoderm outside and endoderm inside . These are the primary germ layers.
The cavity of the primary intestine is formed in the gastrula - gastrocel , which communicates with the external environment through blastopore . Due to the movement of the center of gravity towards the animal pole, the embryo turns 180° with the blastopore upward and continues to float in the water.
Later, the embryo elongates. It is released from the primary endoderm in the dorsal direction. chordal plate, and in the dorsolateral two mesodermal plates. From the primary ectoderm along the midline of the body stands out nervous a plate consisting of higher cells than the rest of the ectoderm. The neural plate is detached from the ectoderm and plunges under it, first turning into philtrum , and then in neural tube , the rest of the skin ectoderm closes over the neural tube. Simultaneously with the formation of the neural tube, the notochordal plate is transformed into a round cellular cord - chord , the mesodermal plates curl into hollow tubes lying between the notochord and the cutaneous ectoderm, and the remaining endoderm closes into secondary colon . This creates a complex of axial organs that characterize the type of chordate animal.
The mesoderm is metamerically (from the head and tail of the embryo) divided into segments, and no segmentation occurs in the tail of the embryo. In addition, the first two segments develop independently, reproducing the ancient three-segmented larval form of skullless - Dipleurula . Each segment of the mesoderm, excluding the first two (“ancient”) segments, grows in the dorsoventral direction and is divided into three parts: somite (dorsally), splanchnotoma (ventrally) and segmental leg between them.
Somites differentiate into dermatome – skin sheet (lateral), sclerotome – skeletal rudiment (central) and myotome – muscle leaf (remnant after isolation of the first two). Skeletal (somatic) muscles subsequently develop from the myotome. The splanchnotome splits into two leaves: visceral (internal) and parietal (parietal), between them there is a secondary body cavity - in general . From both leaves of the splanchnotome, a network-shaped tissue stands out - mesenchyme , which is also formed from the sclerotome and dermotome of the somite. Mesenchyme (embryonic connective tissue) fills the entire space between the three germ layers. From the remaining part of both layers of the splanchnotome, the lining of the coelom arises - mesothelium . Finally, the segmental stalk is transformed into nephrogonotome – epithelial lining of the excretory system and the germ of the reproductive system.
The period of differentiation of tissues and organs ends the larval period of development of the lancelet, which lasts about three months, and a sexually mature animal emerges from the larva.
- In contact with 0
- Google+ 0
- OK 0
- Facebook 0






