Chained inheritance - inheritance of traits whose genes are localized on one chromosome. The strength of linkage between genes depends on the distance between them: the further the genes are located from each other, the higher the frequency of crossing over and vice versa. Along with traits that are inherited independently, there must also be those that are inherited linked to each other, since they are determined by genes located on the same chromosome. Such genes form clutch group. The number of linkage groups in organisms of a certain species is equal to the number of chromosomes in the haploid set (for example, in Drosophila 1 pair = 4, in humans 1 pair = 23).
Full grip- a type of linked inheritance in which the genes of the analyzed traits are located so close to each other that crossing over between them becomes impossible.
Incomplete clutch- a type of linked inheritance in which the genes of the analyzed traits are located at a certain distance from each other, which makes crossing over between them possible.
(Crossover gametes- gametes in the process of formation of which crossing over occurred. As a rule, crossover gametes make up a small part of the total number of gametes.
Crossing over- exchange of sections of homologous chromosomes during cell division, mainly in the prophase of the first meiotic division, sometimes in mitosis. The experiments of T. Morgan, K. Bridges and A. Sturtevant showed that there is no absolutely complete linkage of genes, in which genes would always be transmitted together. The probability that two genes located on the same chromosome will not separate during meiosis ranges from 1 to 0.5. In nature, incomplete linkage predominates, caused by the intersection of homologous chromosomes and gene recombination. The cytological picture of crossing over was first described by the Danish scientist F. Janssens.
Crossing over occurs only when the genes are in a heterozygous state (AB / av). If the genes are in a homozygous state (AB / AB or aB / aB), the exchange of identical sections does not give new combinations of genes in gametes and in the generation. The frequency (percentage) of crossing over between genes depends on the distance between them: the further they are located from each other, the more often crossing over occurs. T. Morgan proposed measuring the distance between genes by crossing over as a percentage, using the formula:
N1/N2 X 100 = % crossing over,
where N1 is the total number of individuals in F;
N2 is the total number of crossover individuals.
The chromosome segment on which 1% crossing over occurs is equal to one morganide (a conventional measure of the distance between genes). Crossing over frequency is used to determine the relative position of genes and the distance between them. New technologies are used to construct a human genetic map; in addition, cytogenetic maps of chromosomes have been constructed.
There are several types of crossing over: double, multiple (complex), irregular, uneven.
Crossing over leads to a new combination of genes and causes a change in phenotype. In addition, along with mutations, it is an important factor in the evolution of organisms.)
The result of T. Morgan's research was his creation of the chromosomal theory of heredity:
· genes are located on chromosomes; different chromosomes contain different numbers of genes; the set of genes of each of the non-homologous chromosomes is unique;
· each gene has a specific location (locus) on the chromosome; allelic genes are located in identical loci of homologous chromosomes;
· genes are located on chromosomes in a specific linear sequence;
· genes localized on the same chromosome are inherited together, forming a linkage group; the number of linkage groups is equal to the haploid set of chromosomes and is constant for each type of organism;
· gene linkage can be disrupted during crossing over, which leads to the formation of recombinant chromosomes; the frequency of crossing over depends on the distance between genes: the greater the distance, the greater the magnitude of crossing over;
· Each species has a unique set of chromosomes - a karyotype.
Inheritance of sex and sex-linked traits. Sex chromosomes and their role in sex determination. Inheritance of sex. The sex of an individual is a complex trait, formed both by the action of genes and developmental conditions. Humans have one of 23 pairs of chromosomes - sex chromosomes, designated X and Y. Women are the homogametic sex, i.e. have two X chromosomes, one received from the mother and the other from the father. Males are a heterogametic sex, have one X and one Y chromosome, with X transmitted from the mother and Y from the father. Note that the heterogametic sex is not always necessarily male; for example, in birds they are females, while males are homogametic. There are other mechanisms of sex determination. Thus, in a number of insects the Y chromosome is absent. In this case, one of the sexes develops in the presence of two X chromosomes, and the other - in the presence of one X chromosome. In some insects, sex is determined by the ratio of the number of autosomes and sex chromosomes. In a number of animals the so-called redetermination of sex, when, depending on environmental factors, the zygote develops into either a female or a male. The development of sex in plants has the same diverse genetic mechanisms as in animals.
Traits linked to the X chromosome. If a gene is located on the sex chromosome (it is called sex-linked), then its manifestation in descendants follows different rules than for autosomal genes. Let's look at the genes located on the X chromosome. A daughter inherits two X chromosomes: one from her mother and one from her father. The son has only one X chromosome - from the mother; He receives a Y chromosome from his father. Therefore, the father passes on the genes on his X chromosome only to his daughter, but the son cannot receive them. Since the X chromosome is more “rich” in genes compared to the Y chromosome, in this sense the daughter is genetically more similar to the father than the son; the son is more similar to his mother than to his father.
One of the historically best known sex-linked characteristics in humans is hemophilia, which leads to severe bleeding from the slightest cuts and extensive hematomas from bruises. It is caused by a recessive defective allele 0, which blocks the synthesis of a protein necessary for blood clotting. The gene for this protein is localized on the X chromosome. A heterozygous woman +0 (+ means the normal active allele, dominant to the hemophilia 0 allele) does not develop hemophilia, and neither do her daughters, unless the father has this pathology. However, her son may receive the 0 allele and then develop hemophilia. Recessive diseases caused by genes on the X chromosome affect women much less frequently than men, since in them the disease manifests itself only in cases of homozygosity - the presence of a recessive allele in each of the two homologous X chromosomes; males are affected whenever their only X chromosome carries the defective allele.
Linkage to the Y chromosome.Information about the genes located on the Y chromosome is very scarce. It is assumed that it practically does not carry genes that determine the synthesis of proteins necessary for the functioning of the cell. But it plays a key role in the development of the male phenotype. The absence of a Y chromosome in the presence of only one X chromosome leads to the so-called. Turner syndrome: the development of a female phenotype with poorly developed primary and secondary sexual characteristics and other deviations from the norm. There are men with an additional Y chromosome (XYY); They are tall, aggressive and often have abnormal behavior. Several genes have been identified on the Y chromosome that are responsible for regulating the synthesis of specific enzymes and hormones, and disturbances in them lead to pathologies of sexual development. There are a number of morphological characters that are believed to be determined by genes on the Y chromosome; among them is the development of ear hair. Characteristics of this kind are transmitted only through the male line: from father to son.
Genetic determination of sex, determined by the set of sex chromosomes, supports equal reproduction of females and males. Indeed, female eggs contain only the X chromosome, since women have the XX genotype on their sex chromosomes. The genotype of men is XY, and therefore the birth of a girl or boy in each specific case is determined by whether the sperm carries an X or Y chromosome. Since during the process of meiosis, chromosomes have an equal chance of getting into a gamete, half of the gametes produced by male individuals contain an X chromosome, and half contain a Y chromosome. Therefore, half of the offspring are expected to be of one sex, and half of the other.
It should be emphasized that it is impossible to predict in advance the birth of a boy or a girl, since it is impossible to predict which male reproductive cell will participate in the fertilization of the egg: carrying the X or Y chromosome. Therefore, the presence of more or fewer boys in a family is a matter of chance.
The law of independent distribution of traits (Mendel's third law) is violated if the genes that determine different traits are located on the same chromosome. Such genes are usually inherited together, i.e. chained inheritance. The phenomenon of linked inheritance was studied by Thomas Morgan and his associates and is therefore called Morgan's law.
T. Morgan's law can be formulated as follows: genes located on the same chromosome form a linkage group and are often inherited together, while the frequency of joint inheritance depends on the distance between genes (the closer, the more often).
The reason why linked inheritance is disrupted is crossing over, which occurs in meiosis during the conjugation of chromosomes. In this case, homologous chromosomes exchange their sections, and thus previously linked genes can end up on different homologous chromosomes, which determines the independent distribution of traits.
For example, gene A is linked to gene B (AB), and the homologous chromosome contains recessive alleles of the corresponding genes (ab). If, during the process of crossing over, homologous chromosomes almost never exchange sections so that one gene moves to another chromosome, while the other remains in the same one, then such an organism forms gametes of only two types: AB (50%) and ab (50%). If an exchange of the corresponding sections occurs, then a certain percentage of the gametes will contain the Ab and aB genes. Usually their percentage is less than with independent distribution of genes (when A and B are on different chromosomes). If, with an independent distribution of all types of gametes (AB, ab, Ab, aB), there will be 25% each, then in the case of linked inheritance of gametes Ab and aB there will be less. The fewer there are, the closer the genes are located to each other on the chromosome.
Sex-linked inheritance is especially distinguished when the gene under study is located on the sex (usually X) chromosome. In this case, the inheritance of one trait is studied, and the second is gender. If an inherited trait is sex-linked, then it is inherited differently during reciprocal crosses (when the trait is first possessed by the female parent, then by the male).
If the mother has the aa genotype, and the father exhibits a dominant trait (there is definitely one gene A), then in the case of sex linkage, all daughters will have a dominant trait (in any case, they will receive his only X chromosome from the father, and all sons will have a recessive one) (from the father one gets the Y chromosome, which does not contain the corresponding gene, and from the mother, in any case, gene a.) If the trait were not sex-linked, then among both sexes of children there could be owners of a dominant trait.
When the genes under study are linked in an autosome, such linkage is called autosomal. Linkage is called complete if the parental combinations of alleles are not disrupted from generation to generation. This happens very rarely. Usually, incomplete linked inheritance is observed, which violates both Mendel's third law and Morgan's law (in its abbreviated form: genes located on the same chromosome are inherited together).
Genes on a chromosome are arranged linearly. The distance between them is measured in centimorgans (cm). 1 cm corresponds to the presence of 1% of crossover gametes. By conducting various crosses and statistically analyzing the descendants, scientists identify linked genes, as well as the distance between them. Based on the data obtained, genetic maps are constructed, which reflect the localization of genes on chromosomes.
Linked inheritance is the phenomenon of correlated inheritance of certain states of genes located on the same chromosome.
There is no complete correlation due to meiotic crossing over, since linked genes can disperse into different gametes. Crossing over is observed in the form of unlinking in the offspring of those gene alleles and, accordingly, states of traits that were linked in the parents.
Observations made by Thomas Morgan showed that the probability of crossing over between different pairs of genes varied, and the idea came to create gene maps based on the frequencies of crossing over between different genes. The first gene map was constructed by Morgan's student, Alfred Sturtevant, in 1913 using material from Drosophila melanogaster.
The distance between genes located on the same chromosome is determined by the percentage of crossing over between them and is directly proportional to it. The unit of distance is taken to be 1% crossing over (1 morganid or 1 centimorganid). The farther genes are from each other on a chromosome, the more often crossing over will occur between them. The maximum distance between genes located on the same chromosome can be 49 centimorganides.
Linked features
Linked traits are traits that are controlled by genes located on the same chromosome. Naturally, they are transmitted together in cases of complete coupling.
Morgan's Law
Linked genes localized on the same chromosome are inherited together and do not exhibit independent distribution
Crossing over
However, homologous chromosomes can cross over (crossing over or decussation) and exchange homologous regions. In this case, the genes of one chromosome are transferred to another, homologous to it. The closer the genes are located to each other on a chromosome, the stronger the linkage between them and the less often their divergence occurs during crossing over, and, conversely, the farther the genes are from each other, the weaker the linkage between them and the more often its disruption is possible.
The number of different types of gametes will depend on the frequency of crossing over or the distance between the genes analyzed. The distance between genes is calculated in morganids: one unit of distance between genes located on the same chromosome corresponds to 1% crossing over. This relationship between distances and crossing-over frequency can be traced only up to 50 morganids.
27. Chromosomal theory of heredity.
The chromosomal theory of heredity is a theory according to which chromosomes contained in the cell nucleus are carriers of genes and represent the material basis of heredity, that is, the continuity of the properties of organisms in a number of generations is determined by the continuity of their chromosomes. The chromosomal theory of heredity arose at the beginning of the 20th century. based on cell theory and was used to study the hereditary properties of organisms through hybridological analysis.
The founder of the chromosome theory, Thomas Gent Morgan, American geneticist, Nobel laureate. Morgan and his students found that:
– each gene has a specific locus (location) on the chromosome;
– genes on a chromosome are located in a certain sequence;
– the most closely located genes on one chromosome are linked, therefore they are inherited predominantly together;
– groups of genes located on the same chromosome form linkage groups;
– the number of linkage groups is equal to the haploid set of chromosomes in homogametic individuals and n+1 in heterogametic individuals;
– an exchange of sections (crossing over) can occur between homologous chromosomes; as a result of crossing over, gametes arise whose chromosomes contain new combinations of genes;
– the frequency (in%) of crossing over between non-allelic genes is proportional to the distance between them;
– the set of chromosomes in cells of a given type (karyotype) is a characteristic feature of the species;
– the frequency of crossing over between homologous chromosomes depends on the distance between genes localized on the same chromosome. The greater this distance, the higher the crossing over frequency. The unit of distance between genes is taken to be 1 morganid (1% crossing over) or the percentage of occurrence of crossover individuals. If this value is 10 morganids, it can be stated that the frequency of chromosome crossover at the locations of these genes is 10% and that new genetic combinations will be identified in 10% of the offspring.
To clarify the nature of the location of genes on chromosomes and determine the frequency of crossing over between them, genetic maps are constructed. The map reflects the order of genes on a chromosome and the distance between genes on the same chromosome. These conclusions of Morgan and his colleagues were called the chromosomal theory of heredity. The most important consequences of this theory are modern ideas about the gene as a functional unit of heredity, its divisibility and ability to interact with other genes.
The formation of the chromosome theory was facilitated by data obtained from the study of the genetics of sex, when differences in the set of chromosomes were established in organisms of different sexes.
In this lesson, we will look at a couple of genes whose inheritance does not obey Mendel’s laws, learn what crossing over and linked inheritance of genes are, and talk about their features.
The formation of these gametes is equal, which formed the basis for the discovery of the third for-co-on of Mende -la - for the un-know-si-my splitting of the signs. However, in the course of a number of experiments, scientists have learned that there are such pairs of signs, in which two types of gametes are found more often than the other two types of gametes.
Let's look at this using a specific example (Fig. 2).

Rice. 2. Drosophila fly ()
In small fruit flies, dro-zo-phil - from-the-love-object-of-ge-ne-ti-kov - gene A from-ve-cha-et for gray body, recessive gene A - for the black color of the body, dominant gene B - for the development of long wings, and recessive the strong gene in - for undeveloped wings, that is, the wings remain in the same state.
When crossing two ge-te-ro-zy-goths, which have a gray body, long wings and AaBb gene types, in the first place generations receive three bodies with a gray body and long wings, as well as one organism with a black body and -cha-precise wings (Fig. 3).

Rice. 3. Crossing of heterozygotes AaBb ()
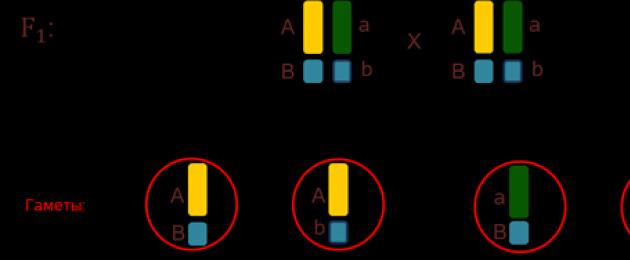
In the future, pay attention to the fact that the signs in the or-ga-bottom are significant pain -sher than the chro-mo-some, in which these signs are lo-ka-li-zo-va-ny. They concluded that, apparently, in one chro-mo-so-me there is a large number of genes. Genes located in one chro-mo-so-me form a single group - clutch, and next most often together (Fig. 4).

Rice. 4. Linked genes ()
This pattern was established by the American gene-ne-ti-k To-ma-s Gen-Mor-ga-n in the na-cha- le 20s last century. Consequently, if the genes lie in the same chromosome, it means that they will be inherited together (Fig. 5).

Rice. 5. Chained inheritance ()
D - gene denoting the gray body; d - black body; F - long wings; f - for-precise wings.
As a result of that kind of cross-breeding, we have three or-ga-niz-ma, having a gray body and a long new wings, and one or-ga-ism, having a black body and very precise wings.
Ge-ne-ti-ki pay attention to the fact that in the offspring of such flies the or-ga-bottoms with a gray body still appear and behind-the-precise wings, as well as with a black body and long wings. An explanation for this was found during the study of cell development - this process is called “meiosis”.
In pro-phase 1, there is a phenomenon of con-jugation or convergence of chromosomes, for which it may follow cross-sin-go-ver- exchange of parts of homologous chromosomes (Fig. 6).

Rice. 6. Cross-syn-go-vera process ()
In the re-zul-ta-th e-ra-zu-yut-sya cross-so-faithful ga-me-you. The or-ga-low-we, which are involved in the result of the fusion of such cross-sovere gametes, are called “ re-com-bi-nant-nye or-ga-niz-we.” Since cross-syn-go-ver does not occur after every con-jugation, then the number of cross-syn-go-ver-gametes is significant but less than the number of necros-verified gametes, the ratio is about 20 to 80%.
During the ex-peri-men-ta To-ma-su Gent Mor-ga-nu managed to prove that often cross-sin-go-ve-ra between the gene -mi directly about the distance between them in chro-mo-so-me, that is, we can say that the further the genes are from each other in chromosomal, the more often cross-syn-go-ver occurs between them .
This is the discovery of pos-vo-li-lo la-bo-ra-to-rii To-ma-sa Mor-ga-na to develop a method, pos-la-yu-schy build chro-mo-some maps, that is, indicate the location of genes in the chro-mo-some for different organisms -niz-mov.
Chro-mo-some maps have been created practically for all agriculturally important animals and plants, work in this right-of-line still continues, although the ge-ne-ti-ki is already using not only the me-home of the gi-bri- di-za-tions, but also others have access to them from time to time.
Bibliography
- Mamontov S.G., Zakharov V.B., Agafonova I.B., Sonin N.I. Biology. General patterns. - Bustard, 2009.
- Ponomareva I.N., Kornilova O.A., Chernova N.M. Fundamentals of general biology. 9th grade: Textbook for 9th grade students of general education institutions / Ed. prof. I.N. Ponomareva. - 2nd ed., revised. - M.: Ventana-Graf, 2005
- Pasechnik V.V., Kamensky A.A., Kriksunov E.A. Biology. Introduction to general biology and ecology: Textbook for grade 9, 3rd ed., stereotype. - M.: Bustard, 2002.
- Polnaja-jenciklopedija.ru ().
- Cribs.me().
- Biorepet-ufa.ru ().
Homework
- Which pairs of genes do not obey Mendel's laws?
- What is the pattern of genes lying on the same chromosome?
- What is crossing over?
- In contact with 0
- Google+ 0
- OK 0
- Facebook 0






