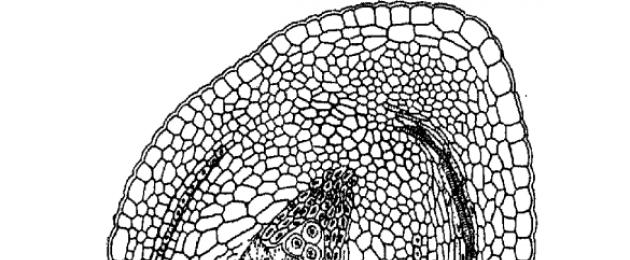
Antes de pasar a la estructura del óvulo, consideremos sus principales tipos morfológicos (Fig.25), establecidos por el botánico francés C. Mirbel en 1829.
En muchas plantas con flores, el micropilo del óvulo está en el mismo eje que el aquenio (funículo) y, por tanto, el óvulo se sitúa perpendicular a la placenta. Un óvulo tan radialmente simétrico se llamaba ortotrópico (del griego orthos - recto, regular y tropos - giro, dirección) o recto. (Figura 25). A menudo se le llama atrópico (del griego a, partícula de negación). Los óvulos ortotrópicos son característicos de Chloranthaceae, Savuraceae, Pepperaceae, Hornwort, Sycamore, Myricaceae, Walnutaceae, Vallisneria, Eelgrass, Pondweed, Xyrisaceae, Eriocaulaceae, Restiaceae y algunos otros géneros y familias.
, 
Mucho más comunes son los óvulos que están doblados hacia abajo (hacia la placenta) 180° y, como resultado, se adhieren de lado al pedúnculo alargado. Por tanto, su micropilo mira hacia la placenta y está situado cerca de la base del óvulo. Estos óvulos se denominan anatrópicos (del griego anátropo - invertido, invertido) o invertidos (Fig. 25, 26). Los óvulos anatrópicos son característicos de la mayoría de las plantas con flores, tanto dicotiledóneas como monocotiledóneas. El famoso botánico checo F. Netolitsky (1926) llegó a la conclusión de que el tipo anatrópico es el original y el ortotrópico es el derivado, lo que concuerda bastante con las ideas modernas.
Junto con los óvulos anatrópicos y ortotrópicos, existen varios tipos más "arquitectónicos", que, sin embargo, son menos comunes. En primer lugar, debemos mencionar los hemitrópicos (del griego hemi - mitad, o, más precisamente, "mitad"), o óvulos medio invertidos, girados sólo 90° con respecto a la placenta. Son característicos, por ejemplo, de la casuarina, de algunas prímulas, de algunas borrajas, de las Lamiáceas, de las Solanáceas y de las Noricháceas, de las Malpighiáceas, de algunas Liliáceas y de muchas otras plantas. Los óvulos hemitrópicos son un tipo intermedio entre anatrópicos y ortotrópicos (Fig. 25).
En los casos en que el óvulo, debido al mayor crecimiento de un lado, se curva, se le llama campilotrópico (del griego kampylos - curvo) o semi-doblado. A diferencia de los tipos anatrópico y hemitrópico, aquí el tegumento e incluso el megasporangio (nucellus) están curvados. Este tipo de óvulo es característico de muchos representantes del orden Cloveaceae, alcaparras, algunas legumbres, Philesiaceae, algunos cereales y varios otros grupos. En la mayoría de los casos, los óvulos campilotrópicos surgen de anatrópicos, pero es posible que en ocasiones también puedan surgir de ortotrópicos.
En algunos casos, el óvulo está doblado por la mitad de tal manera que en un corte longitudinal su megasporangio tiene forma de herradura. Este es un óvulo anfitrópico (del griego amphi - ambos). Se encuentra en Leitneria, Crossosoma, algunas acanthaceae y algunas otras plantas (Fig. 27).

Entre todos estos tipos existen formas intermedias, por lo que muchas veces es difícil determinar qué tipo de óvulo tiene una planta en particular. Sin embargo, la identificación de estos tipos resultó muy útil, ya que tienen un cierto significado sistemático.
Preguntas para el coloquio sobre el tema.
para estudiantes de 2do año de biología
¿Qué es un óvulo? Enumere sus componentes en Gimnospermas.
Óvulo, o óvulo(lat. óvulo) - la formación en las plantas con semillas a partir de la cual (generalmente después de la fertilización) se desarrolla la semilla. Es el esporangio femenino (megasporangio) de las plantas con semillas. En gimnospermas, en la superficie de las escamas de las semillas en conos femeninos, ubicados abiertamente en la axila del megasporofilo. En la parte central del óvulo (nucellus), se forman cuatro megasporas como resultado de la meiosis de la célula madre de la espora, luego tres de ellas mueren y se forma un gametofito femenino a partir de una megaspora. En las gimnospermas, a veces se le llama endospermo porque almacena nutrientes en la semilla madura.
S. consta de una nucel (parte central que contiene un megasporocito), uno o dos tegumentos (tegumentos) y un aquenio (funicular). En la parte superior de S. el tegumento generalmente no se cierra, dejando una abertura estrecha: el micropilo.
¿Qué es la nucela? ¿Cómo ocurre la formación y desarrollo del óvulo?
Nuecesmillús(del latín nucella - nuez), la parte central (núcleo) del óvulo de las plantas con semillas; homólogo megasporangios helechos. Incluye tejido nutritivo y un saco embrionario cubierto de membrana con una pequeña abertura llamada micropilem.

Teorías sobre el origen del tegumento del óvulo.
telomnaya
El tegumento es el resultado de la fusión de cuerpos periféricos inicialmente vegetativos alrededor de un cuerpo portador de esporas. Esta teoría es consistente con los hallazgos paleobotánicos que se relacionan con las protogimnospermas y las gimnospermas extintas: los helechos con semillas.
Sinangial
Según esta hipótesis, propuesta por primera vez por la paleobotánica inglesa Margarita Benson (1908), el tegumento es un anillo de esporangios esterilizados, fusionados y fusionados que rodean el megasporangio de funcionamiento central, y el micropilo corresponde al espacio original entre las puntas de los esporangios. . En otras palabras, el óvulo es en realidad un sinangio en el que todos los esporangios excepto uno han sido esterilizados y han formado el tegumento de un único esporangio fértil. Una buena confirmación de la hipótesis "sinangial" son los óvulos primitivos de los helechos, que a menudo conservan rastros muy claros de su origen sinangial. Los óvulos de varios helechos tenían tegumentos segmentados con un haz vascular en cada segmento (cámara).
Polos micropilar y chalazal del óvulo.
En el primordio de la semilla, la chalaza se ubica frente al micropilo, abriendo el tegumento. La chalaza es el tejido donde se conectan el tegumento y la nucela.
Los dos polos del embrión corresponden a los polos de la raíz y del brote.
En el embrión: hipocótilo y cotiledones.
Características del óvulo como megasporangio modificado de plantas con semillas en comparación con el megasporangio de plantas de esporas superiores.
La semilla es un nuevo tipo de diáspora.
Proceso de megasporogénesis
Hay 2 óvulos en las escamas de los conos femeninos. El óvulo consta de una nucela (núcleo) y un tegumento (cubierta). En la parte superior queda un pequeño agujero: el micropilo (paso del polen). A finales de la primavera, las escamas del cono femenino se abren y el polen cae a través del micrópilo hacia el interior de la nucela. Después de esto, las escamas de las semillas se comprimen, formando protección para los óvulos. Durante la polinización, no hay gametos masculinos en el grano de polvo y el prótalo con arquegonia aún no está desarrollado en el óvulo. Un mes después de la polinización, una de las células de la nucela comienza a dividirse por meiosis. Como resultado, se forman 4 células haploides (n): megasporas. 3 mueren y 1 se convierte en un brote (n). El gametofito femenino es un talo multicelular incoloro. ^ Hay muchas sustancias de almacenamiento en el tejido. 14-15 meses después de la polinización, se forman dos arquegonios en el gametofito. El arquegonio consta de un óvulo grande con un núcleo grande, encima del cual se encuentra una célula del túbulo ventral que desaparece temprano y un cuello de ocho células pequeñas. En este momento, el tubo polínico crece muy lentamente dentro de la nucela. Después de que el tubo polínico con dos espermatozoides llega a la arquegonia, uno de los espermatozoides se fusiona con el óvulo y el otro muere. Un embrión se desarrolla a partir de un cigoto. El óvulo se convierte en semilla. El cuerpo vegetativo del gametofito femenino se convierte en el endospermo primario (n). Se forma una cáscara transparente a partir del núcleo y una cáscara leñosa a partir del tegumento. Así, en la semilla se pueden distinguir partes de 3 generaciones:
Piel leñosa y transparente – esporofito viejo (2n)
Endospermo – gametofito (n)
Embrión – nuevo esporofito (2n)
Desarrollo típico del gametofito femenino de las Gimnospermas.

¿Qué estructuras son homólogas a las anteras y a los granos de polen de las gimnospermas?
Antera – microsporangio, mota de polvo – grano de polen

10. 2 variantes de la estructura de un gametofito masculino maduro, sus elementos celulares.
11. El futuro destino del gametofito masculino de las gimnospermas.
12. ¿Un grano de polen siempre deja la antera madura? ¿Qué opción es más progresista?
13. Polinización (definición)
En las plantas, el polen se transfiere desde las anteras al estigma (en las plantas con flores) o al óvulo (en las gimnospermas). Después de O., a partir de una mota de polvo se desarrolla un tubo polínico, cuyo borde crece hacia el ovario y es liberado por el marido. células sexuales (esperma) al óvulo ubicado en el óvulo, donde se produce la fertilización y el desarrollo del embrión.
14. Fertilización del tipo cícada, sus características.
Tras la polinización, los óvulos comienzan a agrandarse y pronto alcanzan el tamaño de una semilla, aunque en ellos aún no se ha producido la fecundación. Este período, desde la polinización hasta la fertilización, es muy largo y suele durar seis meses (por ejemplo, en las cícadas, la polinización ocurre en diciembre - enero y la fertilización, en mayo - junio).

Germinan las microsporas que ingresan a la cámara de polen con una gota de líquido polinizador. Al mismo tiempo, la exina estalla y la célula de haustorio crece a través del espacio, estirando el intestino (Fig. 168, 13). Penetra en la pared de la cámara de polen y chupa nutrientes del tejido de la nucela (Fig. 168, 13). En este momento, la célula generativa se divide en dos y una de las células resultantes, la espermatogénica, comienza a crecer rápidamente. En él, se forman gametos masculinos, espermatozoides, no inmediatamente, sino después de varios meses (Fig. 168, 15).
En el momento de la fertilización, la célula espermatogénica expandida se encuentra muy cerca de la entrada a la arquegonia. Los espermatozoides liberados sólo tienen que "nadar" en el líquido que se derramó con ellos desde la célula espermatogénica, a poca distancia del arquegonio, en el que el contenido del espermatozoide se fusiona con el óvulo (Fig. 168, 17). ).
Así, en las cícadas se combinan dos mecanismos en un solo proceso, uno de los cuales, la formación de un espermatozoide móvil, es característico de ancestros lejanos fertilizados con ayuda de agua, y el segundo, la formación de un tubo polínico (un tubo polínico en expansión). célula espermatogénica) - es típica de todas las demás plantas que se encuentran más arriba en la “escalera evolutiva” » plantas con semillas.
15. Sifonogamia
(fertilización tipo pino) – fertilización mediante un tubo polínico; el proceso sexual tiene lugar dentro de los óvulos y no depende de la presencia de humedad. La función de producir gametos masculinos la llevan a cabo células especiales.
16. ¿En qué se diferencian fundamentalmente estas opciones de fertilización en las gimnospermas?
17. ¿Qué células (o sus características) en el primer y segundo caso pueden llamarse atávicas?
La semilla se desarrolla en la superficie de la escama de la semilla. Es una estructura multicelular que combina tejido de almacenamiento: endospermo, embrión y una cubierta protectora especial (cubierta de la semilla). Antes de la fertilización, la parte central del óvulo contiene una nucela, que es reemplazada gradualmente por el endospermo. El endospermo es haploide y se forma a partir de los tejidos del gametofito femenino.
Ud. cícadas Y gingko capa exterior de la cubierta de la semilla ( sarcotesta) suave y carnosa, de capa media ( esclerotest) es dura y la capa interna (endotesta) es transparente en el momento en que la semilla madura. Las semillas son dispersadas por varios animales, que comen la sarcotesta sin dañar la esclerotesta.
Ud. tejo Y podocarpo las semillas están rodeadas de carnoso aril- escamas muy modificadas de un cono femenino. Los arilos suculentos y de colores brillantes atraen a los pájaros, que esparcen las semillas de estas coníferas. Los arilos de muchas especies de podocarpus también son comestibles para los humanos.
19. ¿A partir de qué estructuras del óvulo se desarrollan las partes correspondientes de la semilla?
20. ¿Las estructuras de qué etapas del ciclo de vida incluye una semilla?
22. Compara la estructura del ginkgo y las semillas de pino. ¿Cuáles son los signos del primitivismo del primero?
El péndulo desarrollado característico de los pinos degenera cuando el embrión está completamente desarrollado. Una semilla de pino está formada por un embrión, una testa y un megagametofito, que es una reserva de nutrientes.
En las cícadas y el ginkgo, la capa exterior de la cubierta de la semilla (sarcotesta) es suave y carnosa, la capa intermedia (esclerotesta) es dura y la capa interior (endotesta) es transparente en el momento en que la semilla madura.
|
Los óvulos de ginkgo son platispérmicos. |
||||
|
Las estructuras adicionales se agrupan bajo el término angiospermización. |
||||
|
La cámara de polen es unitaria. |
||||
|
Fertilización después de la abscisión. |
||||
23. ¿Cuál es el endospermo primario de una semilla? ¿De qué viene su nombre?
El endospermo primario de las gimnospermas se forma ANTES DE LA FERTILIZACIÓN a partir de una megaspora y corresponde al gametofito femenino. Las células del endospermo de las gimnospermas son inicialmente haploides y luego, como resultado de la fusión nuclear, se vuelven poliploides.
endospermo secundario- tejido formado en las semillas de la mayoría de las plantas con flores DURANTE LA FERTILIZACIÓN.
24. Ventajas evolutivas de la propagación de semillas.
Las semillas son más viables, gracias a los animales, el viento y el agua pueden transportarse a largas distancias. Hay un suministro de nutrientes, el embrión está protegido por las cubiertas de las semillas. La reproducción no está asociada con el agua.
Placentación y sus tipos.
Ovario y sus tipos.
Estructura y tipos de óvulo.
Características generales del gineceo. Significado y estructura
El gineceo es un conjunto de carpelos (carpelos). En el carpelo se desarrolla un óvulo, dentro del cual se forma un saco embrionario: el gametofito femenino, por lo que el gineceo es el órgano femenino de reproducción sexual. Por lo general, los carpelos crecen juntos en sus bordes, formando un recipiente cerrado: un pistilo. El fruto se desarrolla a partir del pistilo después de la fertilización.
Para la reproducción de las plantas, el gineceo es de primordial importancia porque, después de la fertilización, se convierte en un fruto con semillas. De acuerdo con este significado, diferentes partes tienen diferentes grados de importancia para las plantas. Su parte más importante es el óvulo, que se convierte en semilla, por lo que el gineceo sin óvulos pierde su significado biológico y se considera poco desarrollado.
No existen plantas normales sin óvulos, sin embargo, hay plantas cuyo gineceo está representado únicamente por un óvulo. Le sigue en importancia el estigma, es decir, el órgano que recibe el polen. No existe un verdadero estigma sólo en las gimnospermas, donde, sin embargo, es reemplazado por un tejido laxo que se desarrolla en la apertura del óvulo (micropilar). Menos importante es el estilo, que muchas plantas no tienen (muchas ranunculáceas, agracejo, etc.). Sólo las gimnospermas no tienen un ovario verdadero.
El pistilo consta de un ovario, un estilo y un estigma. Las mismas partes se distinguen también en los carpelos. El ovario transporta los óvulos y actúa como una cámara húmeda, protegiendo a los óvulos de la desecación y de ser comidos por insectos y, en parte, de las fluctuaciones de temperatura. Las semillas se desarrollan a partir de los óvulos después de la fertilización. El estigma del pistilo recibe polen, por lo que a menudo la superficie del estigma es desigual, grumosa y cubierta con un líquido pegajoso. El estilo eleva el estigma. A veces no hay estilo y se forman estigmas sésiles.
La posición del ovario en la flor depende del grado de fusión con el receptáculo y el perianto. Hay ovarios superiores, inferiores y semiinferiores.
El ovario superior se ubica libremente sobre el receptáculo, sin fusionarse con otras partes de la flor. En este caso, partes del perianto se encuentran debajo del ovario, por lo que estas flores también se llaman subpistiladas. Cuando el ovario se fusiona completamente con el receptáculo y las bases de los pétalos y estambres, se forma el ovario inferior. Los tallos florales están unidos al receptáculo encima del ovario, formando una flor suprapistal. En el ovario semiinferior sólo su parte inferior se fusiona con el receptáculo.
Se supone que el ovario inferior está protegido de forma más fiable contra condiciones adversas externas.
El gineceo de diferentes plantas es extremadamente diverso no solo en su composición, número de partes y las circunstancias anteriores, sino también en la forma y tamaños relativos de sus partes. En las gimnospermas, consta de dos o más óvulos que no están cerrados en un ovario; en lugar de un ovario, suelen tener una hoja en forma de escama, en cuya base se asientan (pinos, abetos, abetos, etc.). El resto de plantas con flores siempre tienen el ovario más o menos cerrado, por eso se les llama angiospermas. Además, hay plantas en las que toda la flor está formada únicamente por el gineceo, incluso sin cobertura (sauce).
Actualmente, existen tres tipos de gineceo:
1) Gineceo apocarpo: consta de muchos carpelos independientes, caracterizados por una placentación marginal.
2) Gineceo monocárpico: consta de un solo pistilo con placentación marginal.
3) Gineceo cenócarpo: consta de varios pistilos fusionados. Hay tres variedades de genicea cenocarposa:
a) Sincarpos: varios carpelos fusionados con paredes, placentación central-angular.
b) Paracarpos: lo mismo, pero las paredes de los carpelos no se han conservado. La colocación es en la pared.
c) Lisícarpo: se diferencia del paracarpo por la placentación columnar (central).
El gineceo puede ser simple o complejo. Un gineceo compuesto o apocarposo consta de varios carpelos no fusionados (frambuesa, ranúnculo).
Un gineceo simple puede contener un carpelo, entonces se llama monocarposo, o un pistilo de muchos carpelos fusionados; este es un gineceo cenocarposo. Hay tres subtipos de gineceo cenócarpo según el método de fusión del carpelo: sincárpico, paracarpos y lisícarpo. En un gineceo sincarpo, los carpelos crecen junto con sus superficies laterales formando un ovario multilocular, y los óvulos se ubican más cerca del centro de este ovario (manzano, algodón). El gineceo paracarpos está formado por carpelos con bordes fusionados, por lo que el ovario es unilocular (uvas, grosellas). El gineceo lisicarpo se desarrolla a partir del sincarpo, pero al mismo tiempo las paredes internas del ovario se disuelven y los óvulos se ubican sobre una columna en el centro del ovario (dientes).
La estructura del óvulo antes de la fertilización incluye un aquenio (funículo), una placenta, dos tegumentos y una nucela. La placenta es el lugar donde el tallo de la semilla se une al ovario. La parte interna del óvulo, la nucellus, está cubierta por dos tegumentos: tegumentos, externo e interno. Los tegumentos en el vértice del óvulo no se cierran, formando una abertura microscópica: el conducto polínico o micropilo. La parte de la nucela que mira al micropilo se llama polo micropilar y el lado opuesto se llama chalaza o polo chalazal.
Después de que se forma una megaspora a partir de una célula nucela, crece en el óvulo hacia el saco embrionario, que ocupa la mayor parte, y la nucela permanece en forma de una pequeña capa de pared.
Fig.1 Gineceo
Origen y evolución del gineceo
En el proceso de evolución del gineceo, los carpelos crecen gradualmente juntos y del gineceo apocarposo surge el gineceo cenocarposo (del griego kainos - común). En el gineceo ceenocarposo, las columnas individuales (estilodios) pueden permanecer libres o crecer juntas para formar una columna común (columna compuesta). Hay tres tipos de gineceo cencarpo: sincarpo, paracarpo y lisicarpo.
Sincarpo (del griego syn - juntos) es un gineceo que consta de un número diferente de carpelos cerrados, fusionados por partes laterales. Es bilocular y se caracteriza por el hecho de que los óvulos se ubican a lo largo de las suturas de los carpelos cerrados, es decir. en los ángulos formados por sus partes abdominales (la llamada placentación angular). El gineceo sincarpo generalmente deriva de un gineceo apocarposo con una disposición cíclica (circular) de carpelos, pero en algunos casos también deriva de un gineceo en espiral. Un buen ejemplo de gineceo sincárpico es el del lirio y el tulipán. En las primeras etapas de la evolución del gineceo sincarpo, solo los ovarios de los carpelos crecen juntos y los estilos (estilodios) permanecen libres. Pero gradualmente el proceso de fusión también involucra a las columnas, que eventualmente crecen juntas formando una columna compleja que termina en una cabeza estigmática, que se puede ver en los brezos o en la mayoría de las monocotiledóneas, incluido el lirio.
Mucho más común que el gineceo sincárpico es el gineceo paracarpos (del griego para - cerca). El gineceo paracarpos se caracteriza por la apertura de los carpelos individuales manteniendo la conexión de las áreas marginales de los carpelos vecinos. A diferencia del gineceo sincárpico, el gineceo paracarpos es morfológicamente unilocular y se caracteriza por la llamada placentación parietal o parietal (del latín parietalis - pared, Fig. 19). Ejemplos bien conocidos de gineceo paracarpos son la amapola, el pepino y la calabaza. En muchos casos, el gineceo paracarpo evolucionó a partir del sincarpo. Probablemente esto se debió al hecho de que con la aparición del gineceo sincárpico, la necesidad de cerrar cada carpelo individual, al menos en muchos casos, se vuelve, en esencia, innecesaria. Por tanto, no es de extrañar que el gineceo paracarpos haya surgido de forma independiente y paralela en las más diferentes líneas de evolución de las plantas con flores y caracterice a muchas familias y órdenes enteros. Estructuralmente, el gineceo paracarpo es más económico que el sincarpo: con un gasto relativamente menor en material de construcción, se garantiza una protección y un suministro bastante eficaz de los óvulos y del embrión en desarrollo. Además, los tubos polínicos tienen un acceso más amplio a los óvulos que en el ovario multilocular del gineceo sincarpo.
En muchos otros casos, lo más probable es que el gineceo paracarpo haya evolucionado directamente a partir del apocarpo. Este es probablemente el origen del gineceo paracarpo de algunos géneros africanos de la familia Annonaceae, la mayoría de cuyos representantes se caracterizan por un gineceo apocarpo, o el origen del gineceo paracarpo de la familia Canella. El mismo, según todos los datos, es el origen del gineceo paracarpo de los representantes de las familias Savuraceae y Cactaceae, todo el orden Violaceae y varios órdenes descendientes de él, así como la familia Aquaticaceae entre las monocotiledóneas.
En el gineceo paracarpos suelen crecer placentas, que a menudo incluso se ramifican. En muchos casos, estas placentas agrandadas chocan en el centro del ovario, se fusionan entre sí y la cavidad del gineceo unilocular se divide en cámaras, que son falsos nidos. Las placentas agrandadas se expresan bien en algunas cucurbitáceas, como el pepino y la sandía.
En algunas líneas de desarrollo de dicotiledóneas, por ejemplo, en purslanáceas y prímulas, un gineceo lisícarpo surgió del gineceo sincárpico (del griego lisis - desatar, liberación, disolución). Se trata de un gineceo unilocular, en el que la placentación no es parietal, como en el gineceo de tipo paracarpo, sino la llamada central libre o columnar. La unilocularidad del gineceo lisicarpo se produce en el proceso de evolución (o en la ontogénesis) como consecuencia de la desaparición de los septos (paredes laterales) del ovario sincarpo. En este caso, las partes marginales fusionadas no colapsan ni divergen, y los óvulos continúan posándose sobre ellas. Así, estas partes marginales restantes de los carpelos, junto con las placentas, se convierten en una columna central que se eleva en la cavidad del ovario.
Además de estos tres tipos principales de gineceo cenócarpo, también existe el llamado gineceo pseudomonomérico. Se trata de una versión muy reducida del gineceo sincarpo o paracarpo, en el que sólo un carpelo es fértil y está completamente desarrollado. Los carpelos estériles en un gineceo pseudomonomérico suelen ser tan reducidos que su presencia sólo puede detectarse como resultado de un estudio especial de su estructura anatómica y su desarrollo. Se puede observar un gineceo pseudomonomérico típico en el olmo, la morera, la ortiga, el cáñamo y plantas afines.
En muchas plantas con flores, incluidos los grupos primitivos, el gineceo todavía está libre, no fusionado con las partes circundantes de la flor. Pero en muchas líneas de evolución de las plantas con flores, de forma completamente independiente y en paralelo, se produjo una mayor o menor fusión del gineceo (generalmente cenocárpico) con las partes circundantes de la flor, y como resultado surgió el llamado ovario inferior. Este proceso se produjo de forma gradual y existen todas las formas intermedias entre el ovario superior e inferior. Por tanto, en la morfología de las flores, además de los términos ovario “superior” e “inferior”, también existe el término ovario “semi-inferior”. El origen del ovario inferior es una de las cuestiones difíciles en la morfología evolutiva de las plantas, y se le dedica una gran cantidad de literatura. Para resolver esta cuestión fue de gran importancia el estudio anatómico comparativo de la flor, especialmente de su sistema conductor. La importancia del sistema conductor para resolver diversos tipos de problemas morfológicos se basa en su constancia relativamente mayor (conservadurismo). Los datos sobre la anatomía del sistema conductor de las flores con ovario inferior llevan a la conclusión de que en la gran mayoría de los casos el ovario inferior se produjo como resultado de la fusión de las bases de los sépalos, pétalos y filamentos estaminados con el ovario. El crecimiento de un tubo floral muy ajustado al ovario es bastante natural y está biológicamente justificado. Las diferentes etapas de este crecimiento del tubo floral hasta el ovario se pueden rastrear, por ejemplo, en la familia de los brezos. Sin embargo, como han demostrado los mismos estudios anatómicos comparativos, el origen del ovario inferior puede ser diferente. En algunas familias, como Aizoonaceae, Cactaceae, Santalaceae y familias relacionadas, así como en otras, el ovario no está cubierto de un tubo floral, sino de un receptáculo. El recorrido de los haces vasculares muestra claramente que el ovario parece haberse hundido en el hueco en forma de copa del receptáculo y fusionado con él. Sin embargo, esta segunda forma de formación del ovario inferior se observa sólo en un número muy reducido de familias.
Placentación y sus tipos.
La placentación es el método de disposición de placentas y óvulos (óvulos) en carpelos en los ovarios de las plantas con flores. Las diferencias en la placentación son importantes para comprender la evolución de las plantas con flores.
La ubicación de las placentas, así como la ubicación de los óvulos, varía entre los diferentes grupos de plantas con flores y está estrechamente relacionada con uno u otro tipo de gineceo. Hay dos tipos principales de placentación: sutural o marginal y laminal o superficial.
En el gineceo apocarposo (incluidos los monoméricos), la placentación sutural o marginal se expresa claramente. En este caso, los óvulos se ubican a lo largo de los bordes de los carpelos en dos filas a lo largo de las suturas ventrales.
En el gineceo cencarpnoso (sincarpnoso, paracarpnoso y lisocarpnoso) existen diferentes tipos de placentación sutural. Esto se debe a las peculiaridades de la fusión de carpelos.
El gineceo sincarpo se caracteriza por la fusión de carpelos conduplicados o involutos. Crecen juntos en partes laterales, formando tabiques que separan los nidos de ovario formados, cuyo número corresponde al número de carpelos fusionados. Las suturas ventrales de los carpelos que llevan la placenta aparecen en el centro del ovario. Este tipo de placentación se llama centro-marginal o centro-angular (lirio, campanilla, borraja).
En el gineceo paracarpo, los carpelos no están fusionados por sus superficies laterales, sino por sus bordes, formando un ovario unilocular. La placentación en este tipo de gineceo es similar a una pared o pariental: pared marginal si las placentas están ubicadas en los bordes de los carpelos y pared dorsal si están confinadas a las nervaduras centrales de los carpelos. Es característico de especies de muchas familias y órdenes (brassica, amapola, violeta, sauce, orquídea, etc.).
El gineceo lisicarposo también tiene un ovario unilocular, pero las placentas que contiene están ubicadas en una columna que se eleva desde la parte inferior del ovario. Este tipo de placentación se llama columnar o axial (a veces central libre). La placentación columnar es característica de prímulas y clavos.
Las angiospermas primitivas se caracterizaban por una placentación laminar, que también se encuentra en algunas plantas con flores modernas (magnolia, loto, susak, hornwort, nenúfar). Este tipo de placentación se caracteriza por la unión de los óvulos no a lo largo de la sutura ventral, sino a lo largo de toda la superficie interna del ovario o en ciertos lugares.
El lugar donde se unen los óvulos al ovario se llama placenta. La placenta suele verse como una pequeña hinchazón, crecimiento o protuberancia formada por los tejidos del ovario.
Dependiendo de las características de la unión de los óvulos a la pared del ovario, se distinguen varios tipos de placentación (Fig. 2). La ubicación de los óvulos puede ser parietal o parietal, cuando los óvulos se ubican dentro del ovario a lo largo de sus paredes exteriores o en sus protuberancias. En el caso de que los óvulos se encuentren en la columna central del ovario, divididos en nidos según el número de carpelos, la placentación se denomina axial o axial. Con placentación central libre, los óvulos se desarrollan en una columna central libre, no conectada por tabiques a la pared del ovario. En algunos casos, el único óvulo se encuentra en la base misma de un ovario unilocular. Este tipo de placentación se llama basal. Existen otros tipos de placentación, pero son relativamente raros. La diferencia de placentación es de gran importancia en la taxonomía de las plantas con flores.

Fig. 2.: 1 - pared o parietal, 2 - axial o axial, 3 - central libre, 4 - basal.
Ovario y sus tipos.
Ovario es un término de morfología vegetal; Recipiente hueco cerrado, la parte inferior hinchada del pistilo de una flor bisexual o femenina. El ovario contiene óvulos protegidos de forma segura. Después de la fertilización, el ovario se convierte en un fruto, dentro del cual se encuentran las semillas que se desarrollan a partir de los óvulos.
El ovario actúa como una cámara húmeda que protege a los óvulos de la desecación, las fluctuaciones de temperatura y de ser comidos por insectos.
El ovario y el estigma del pistilo, que sirve para atrapar el polen, están conectados por una columna (si hay varios pistilos en una flor, sus partes superiores estrechas se llaman estilodos).
El ovario puede ser unilocular o multilocular (en este último caso está dividido por tabiques en nidos; a veces los nidos están separados por falsos tabiques).
Según el tipo de ubicación en la flor, los ovarios se denominan:
Ovario superior (libre): unido por su base al receptáculo, sin fusionarse con ninguna parte de la flor (en este caso, la flor se llama subpistilada o peripistótica).
El ovario inferior se encuentra debajo del receptáculo, las partes restantes de la flor están unidas en su ápice (en este caso la flor se llama suprapistal).
Ovario semiinferior: se fusiona con otras partes de la flor, hipantio o receptáculo, pero no en la parte superior, su vértice permanece libre (en este caso la flor se llama semisupapistal).

Fig. 3. tipos de ovarios
Estructura y tipos de óvulo.
Óvulo u óvulo, formación multicelular en las plantas con semillas a partir de la cual se desarrolla la semilla. Las partes principales de S. son la nucela, el tegumento (o tegumentos) y el aquenio. La nucela típicamente surge en forma de tubérculo a partir de las células de la megasporofila (carpelo) de la placenta. El tegumento se forma en forma de cresta anular en la base de la nucela y crece demasiado sobre la nucela en desarrollo, dejando sobre su vértice un canal estrecho: el micropilo o pasaje de polen, debajo del cual en la mayoría de las gimnospermas hay una cámara de polen. El tallo de la semilla (funículo) se conecta. con la placenta. la parte basal desde donde se extiende el tallo de la semilla, llamada chalaza.
En el ovario del pistilo hay pequeñas formaciones: óvulos. Su número oscila entre uno (trigo, ciruela) y varios millones (en los archidae). Las funciones del óvulo son la megasporogénesis (creación de megasporas) y la megagametogénesis (formación del gametofito femenino, proceso de fecundación). El óvulo fertilizado se convierte en una semilla. La placenta es el lugar donde el óvulo se une al carpelo.
Partes del óvulo:
nucellus, núcleo del óvulo;
funículo, el pedúnculo con el que se une el óvulo a la placenta;
tegumentos, cubiertas del óvulo, que forman un canal en el ápice de la nucela;
micropilo, paso del polen;
chalaza, la parte basal del óvulo, donde se fusionan la nucela y los tegumentos;
hilio: el lugar donde el óvulo se une al pedúnculo.

Fig.4 Estructura del óvulo
Tipos de óvulo:
ortotrópico: recto, funículo y micropilo están ubicados en los extremos opuestos del eje del óvulo (trigo sarraceno, nuez);
anatrópico (inverso): la nucela gira 180 con respecto al eje directo del óvulo, como resultado de lo cual el micropilo y el funículo se ubican cerca (angiospermas)
hemitrópico (semi-rotado): el óvulo gira 90, como resultado de lo cual el micropilo y la nucela se ubican en relación con el funículo en un ángulo de 90 (prímulas, norica).
campilotrópico (curvado unilateralmente): la nucela está curvada unilateralmente con un extremo micropilar, respectivamente, el micropilo y el funículo se encuentran cerca (leguminosas, malváceas)
anfitrópico (curvado bilateralmente): la nucela está curvada bilateralmente en forma de herradura, con el micropilo y el funículo ubicados cerca (morera, jara).

Fig.5 Principales tipos de óvulos
6. Desarrollo del óvulo
La megasporogénesis ocurre en la esfera reproductiva femenina, en el gineceo. Morfológicamente, el gineceo está representado por un pistilo (o pistilos). La composición del pistilo incluye: estigma, estilo y ovario. El ovario contiene óvulos (uno o más). El contenido interno del óvulo es una nucela. El tegumento del óvulo está formado por un tegumento doble o único. La nucellus del óvulo contiene una célula archesporial (2n), capaz de dividirse por meiosis (en los sauces y algunas otras plantas el archesporium es multicelular). Como resultado de la meiosis, se forman cuatro megasporas haploides (n) a partir de la célula archesporial (célula madre de las megasporas). Pronto tres de ellos mueren y uno aumenta de tamaño y se divide tres veces mediante mitosis. Como resultado, se forma un saco embrionario de ocho núcleos (gametofito femenino). Tres núcleos, junto con el citoplasma adyacente, forman células antípodas, dos núcleos, un núcleo diploide central; dos núcleos - dos células sinérgicas; un núcleo se convierte en el núcleo del huevo.
Cuando un tubo polínico se acerca al óvulo, lo “siente” de antemano y se prepara para encontrarse con él. Las células compañeras comienzan a secretar sustancias mucosas. Mientras tanto, el tubo polínico crece, venciendo la resistencia de las células de las paredes del ovario. Finalmente llega al micrópilo. Se produce un proceso “dramático”: el tubo polínico perfora (y en el proceso mata) una de las células compañeras. Ambos espermatozoides abandonan el tubo polínico. El destino de la célula vegetativa de un grano de polen no es envidiable: pronto morirá. Es muy difícil observar este proceso, pero aún lo es más entender lo que sucede durante la fecundación.
En agosto de 1898, cuando aún no se conocía la doble fecundación, se celebró en Kiev el Décimo Congreso de Naturalistas y Médicos Rusos. El profesor Sergei Gavrilovich Navashin hizo un informe importante al respecto: ambos espermatozoides contenidos en el grano de polen son necesarios para el desarrollo normal de las semillas de dos especies de la familia de las liliáceas: la azucena (Lilium martagon) y el urogallo (Fritillaria tenella). ¿Por qué Navashin eligió estas plantas en particular? Probablemente porque sus espermatozoides y su saco embrionario son grandes, son fáciles de examinar bajo un microscopio. Navashev fue el primero en afirmar que el proceso de fertilización implica la fusión de dos espermatozoides con dos (!) células del saco embrionario. Pero demos la palabra al propio autor del descubrimiento.
Cada vez que se observó un tubo polínico en contacto con el saco embrionario, también se observaron ambos núcleos sexuales masculinos en el contenido del saco embrionario. Los núcleos masculinos al principio se encuentran cerca uno del otro...
Los núcleos masculinos se separan entonces entre sí, uno penetra en el óvulo y el otro está estrechamente unido a uno de los núcleos polares que aún no se ha fusionado en este momento, es decir, al núcleo hermano del óvulo...
Mientras que el núcleo masculino se adhiere cada vez más estrechamente al núcleo del óvulo, el núcleo polar, copulando con otro núcleo masculino, se desplaza hacia otro núcleo polar, con el que se encuentra en medio del saco embrionario...
Sólo después de la profase de división se fusionan los núcleos..."
Para aclarar la explicación, Sergei Gavrilovich hizo dibujos que gustaron mucho a los botánicos de la capital. Los llevaron para mostrarlos en una reunión de la Academia de San Petersburgo.
¿Por qué los científicos concedieron tanta importancia a este discurso? Se creía que la fertilización en las plantas ocurre de la misma manera que en los animales. Un espermatozoide y un óvulo deberían producir un cigoto a partir del cual se desarrolla una nueva planta. No es que los botánicos no hayan visto dos espermatozoides moviéndose en un tubo polínico hacia el saco embrionario. Se creía que se trataba de algún tipo de anomalía, que una planta "típica" no debería tener dos, sino un espermatozoide, y la fertilización "típica" debería ser la misma tanto en plantas como en animales. Resultó que estas ideas estaban equivocadas. La fertilización de plantas con flores es completamente diferente de la fertilización no solo de animales, sino también de otras plantas. El proceso de fertilización utilizando dos espermatozoides se llama doble fertilización. Inmediatamente después del descubrimiento de Navashin, los científicos se apresuraron a buscar medicamentos. Resultó que muchos ya habían visto la doble fertilización, pero la consideraban un proceso feo que supuestamente no producía semillas normales. Los científicos comenzaron a enviar felicitaciones a Sergei Gavrilovich. Y un científico incluso donó sus viejos preparados, que mostraban claramente una doble fertilización.
Entonces, S.G. Navashin descubrió un fenómeno asombroso. Estaba claro por qué se produce la fertilización del óvulo: para crear un cigoto y, a partir de él, una nueva planta. Pero ¿por qué fertilizar la célula central del saco embrionario? Resultó que es a partir de esta célula que se desarrolla el tejido nutritivo de la semilla, el endospermo. Navashin sugirió que el endospermo no se puede formar sin un núcleo masculino. ¿Cómo podemos probar esta suposición? Después de todo, es imposible eliminar uno de los espermatozoides del tracto polínico sin interrumpir el proceso de fertilización.
Sergei Gavrilovich decidió estudiar plantas que no tienen endospermo. ¿Qué pasa si se altera la fertilización de la célula central? Estas plantas se encuentran en la familia de las orquídeas (Orchidaceae). Las semillas de orquídeas son muy pequeñas y ni siquiera pueden germinar por sí solas (germinan solo con la ayuda de hongos basidiomicetos, formando micorrizas). Navashin vio dos espermatozoides en el tubo polínico de la orquídea. Uno de ellos fecundó el óvulo y el segundo “intentó” fertilizar la célula central con dos núcleos polares. ¡Pero en la célula central los núcleos no se fusionaron entre sí! El proceso de fertilización se vio interrumpido y, naturalmente, no se pudo formar el endospermo. En trabajos posteriores, el científico pudo demostrar que en el girasol (Helianthus annuus) y en algunas otras plantas la fertilización es doble. Después de que Navashin descubriera la doble fertilización en una variedad de plantas, concluyó que la doble fertilización es característica de todas las plantas con flores.
Echemos un vistazo a la estructura del óvulo después de la doble fertilización. Las células de ambos tegumentos tenían dos juegos de cromosomas y los retenían. Ambos conjuntos de estas células pertenecen a la planta madre. Nucellus también porta dos juegos maternos de cromosomas. En el saco embrionario, los sinérgicos murieron y las antípodas tenían cada una un juego de cromosomas. Estos conjuntos también son maternos. El cigoto, formado por la fusión de un óvulo y un espermatozoide, porta dos juegos de cromosomas: uno de la planta paterna y otro de la planta materna. Lo más interesante es que la célula central tiene tres juegos de cromosomas: uno de la planta madre y dos de la planta madre.
Después de la doble fertilización, comienzan varios procesos: el núcleo del endospermo primario se divide para formar el endospermo, el cigoto se desarrolla hasta convertirse en el embrión, los tegumentos se desarrollan hasta convertirse en la cubierta de la semilla y la pared del ovario y las estructuras asociadas forman el fruto. En las primeras etapas de desarrollo, la secuencia de divisiones celulares en embriones dicotiledóneos y monocotiledóneos es similar; en ambos casos se forman cuerpos esféricos. Posteriormente aparece una diferencia: el embrión de las dicotiledóneas tiene dos cotiledones, mientras que el de las monocotiledóneas tiene uno solo. En las semillas de algunos grupos de angiospermas, la nucela se convierte en un tejido de almacenamiento llamado peristermo. Algunas semillas contienen tanto endospermo como peristermo, como en la remolacha (Belta). Sin embargo, en muchas dicotiledóneas y algunas monocotiledóneas, todo o la mayor parte del tejido de reserva es absorbido por el embrión en desarrollo incluso antes de que la semilla entre en estado latente (en guisantes, frijoles, etc.). Los embriones de estas semillas suelen desarrollar cotiledones carnosos llenos de nutrientes. Los principales nutrientes almacenados en las semillas son los carbohidratos, las proteínas y los lípidos. Las semillas de gimnospermas y angiospermas difieren en el origen de estas sustancias. En el primero son producidos por el gametofito femenino, en el segundo por el endospermo, que no es tejido gametofítico ni esporofito.
El desarrollo del óvulo en semilla va acompañado de la transformación del ovario (y a veces de otras partes de la planta) en fruto. Durante este proceso, la pared del ovario (pericarpio o pericarpio) a menudo se espesa y se diferencia en capas separadas: el exocarpio externo (extracarpio), el mesocarpio medio (intercarpio) y el endocarpio interno (intracarpio), generalmente más visible en los carnosos que en los carnosos. en frutos secos.
Así es como se desarrolla el gametofito, la fertilización y la formación de semillas ocurren en una planta con flores "típica", pero el mundo vegetal es muy diverso y en muchas angiospermas el desarrollo se desarrolla de manera diferente, pero no menos interesante y sorprendente. Todos estos métodos de desarrollo ayudan a cada planta a mantener su estrategia en la lucha por la existencia; La ventaja de la doble fertilización no se limita al hecho de que las plantas forman reservas nutricionales en el óvulo sólo bajo la condición de fertilización (sin "desperdiciar" energía en la formación de reservas nutricionales en el óvulo, que nunca será fertilizado, como suele suceder en las gimnospermas), estas diferencias en el desarrollo, que son consecuencia de la adaptación de las plantas a las condiciones del entorno, y crean esa asombrosa variedad de formas de estructura morfológica y fisiológica que observamos en las plantas con flores.
7. Saco embrionario y su desarrollo.
El gametofito femenino de las plantas con flores suele denominarse saco embrionario. El origen de este nombre se remonta a aquellos tiempos en los que aún no estaba clara la naturaleza morfológica del peculiarísimo gametofito femenino de las plantas con flores. Actualmente, ambos términos se utilizan como sinónimos.

Fig.6. Saco embrionario (prótalo femenino) de una planta angiosperma (floreciente) y doble fertilización: 1 - huevo; 2 - células auxiliares; 3 - tubo polínico abierto. Uno de los espermatozoides (4a) se fusiona con el óvulo; 5 - núcleos polares que se fusionan con el segundo espermatozoide (46); 6 - tres células en la parte inferior del saco embrionario (antípodas).
En la mayoría de las plantas con flores (probablemente al menos el 80%), el gametofito femenino se inicia mediante una de las cuatro megasporas tétradas potenciales a través de tres divisiones mitóticas sucesivas. Este tipo de formación del gametofito femenino se denomina monospórica. Su desarrollo se produce de la siguiente manera. Como resultado de la primera división del núcleo de una megaspora en funcionamiento, se forman dos núcleos, que divergen hacia los polos (micropilar y chalazal) de una megaspora fuertemente alargada, y se forma una gran vacuola entre ellos. A continuación, cada uno de estos dos núcleos se divide sincrónicamente dos veces más y como resultado se forman cuatro núcleos en cada polo. Esta es la etapa de ocho núcleos del desarrollo del gametofito femenino. De cada uno de los dos cuádruples polares, un núcleo ingresa a la parte central del saco embrionario. Estos llamados núcleos polares se juntan en la parte central y se fusionan inmediatamente o solo antes de la fertilización y, a veces, no se fusionan en absoluto. Como resultado de la fusión de los núcleos polares, se forma el núcleo de la célula central del gametofito femenino, a menudo llamado núcleo secundario del saco embrionario.
Posteriormente (durante la última mitosis), surgen particiones celulares y la etapa de desarrollo nuclear libre del gametofito femenino pasa a la etapa celular. En el polo micropilar se forman tres células del aparato óvulo, en el polo opuesto (chalazal) hay tres células llamadas antípodas, y entre estos dos grupos de células hay una célula central (que contiene dos núcleos polares libres o un núcleo secundario) . Las tres células del aparato del óvulo se diferencian en un óvulo y dos células, llamadas sinérgicas (del griego synergos, que actúan juntas) o células auxiliares. Ésta es la estructura del llamado gametofito femenino monospórico normal.
El gametofito femenino monospórico normal no da la impresión de ser un modelo completamente desarrollado en el que la economía haya alcanzado el mismo límite que en el caso del gametofito masculino. El hecho de que aquí no se haya alcanzado el límite lo demuestra lo siguiente. En el proceso de evolución de este tipo de gametofito femenino, principal tipo de plantas con flores, aparecieron sacos embrionarios de otros tipos, formados por dos o incluso cuatro megasporas. Hubo una reducción de las divisiones mitóticas a dos o una, y la distribución de los núcleos también cambió. Diferentes combinaciones de estos cambios llevaron al surgimiento de varios tipos de gametofitos femeninos, que se diferencian tanto en el número de núcleos (4, 8, 16), como en el número de grupos celulares y en los núcleos polares (1, 2, 4, 7-14). ), y el número de células en grupos (por ejemplo , el aparato del óvulo puede constar de 1, 2, 3, 5 y 7 células), así como otras características.
La característica principal del tipo de formación monospórica del gametofito femenino es la formación secuencial de particiones celulares durante las divisiones meióticas y la formación de megasporas separadas. La megasporogénesis, que precede a la formación de gametofitos bispóricos (de dos esporas) y tetraspóricos (de cuatro esporas), tiene un carácter completamente diferente. Los cambios en la megasporogénesis consisten en la supresión de la formación de un tabique celular durante la división meiótica, como resultado de lo cual, en lugar de tétradas de megasporas, aparecen formaciones no celulares llamadas cenocitos (del griego koinos - general y kytos - vaso, célula). Aparecen, que luego se desarrollan hasta convertirse en gametofitos femeninos. Hay dos opciones posibles aquí.
En el primer caso, la formación de particiones celulares se suprime sólo durante la segunda división meiótica. Por tanto, ambos núcleos incluidos en la célula viable de la díada participan en la formación del gametofito femenino. Una díada binuclear es un cenocito, producto de la fusión (o más bien, de la no separación) de dos megasporas potenciales. Dado que cada uno de estos cenocitos corresponde en origen a un par de megasporas no separadas, se considera un cenocito de dos esporas y el gametofito femenino que se desarrolla a partir de él se considera bispórico (de dos esporas). El gametofito femenino bispórico se observa en cebollas, escila, lirio de los valles, amarilis, chastuha y algunas otras plantas. El desarrollo de dicho gametofito femenino se produce de forma más abreviada y sólo se requieren dos divisiones mitóticas para formar la etapa de ocho núcleos.
Finalmente, en algunas plantas, como peperomia, olmo, maynik, urogallo, tulipán, azucena, cebolla de ganso, plumbago, adoxa y otras, la formación de particiones celulares se suprime durante ambas divisiones meióticas. Aquí toda la célula madre de la megaspora se transforma en un cenocito de cuatro esporas o tetracenospora. Un cenocito de cuatro esporas corresponde a cuatro megasporas indivisas, por lo que los gametofitos femeninos de este tipo se denominan tetraspóricos (de cuatro esporas). Para formar la etapa de ocho núcleos del gametofito femenino, solo se requiere una división mitótica de los núcleos del cenocito de cuatro esporas.
Según I.D. Romanov, a diferencia del gametofito femenino monospórico, los bi y tetraspóricos se denominan cenospóricos.
El gametofito femenino de las plantas con flores evolucionó a partir del gametofito femenino arquegonial más primitivo de ancestros gimnospermas lejanos a través de la pérdida de las últimas etapas de desarrollo que conducen a la formación de arquegonios. Y en los tipos cenosporicos, como hemos visto, las etapas iniciales también caen. Pero mientras la evolución del gametofito masculino ha llegado a su etapa culminante y todas las posibles etapas iniciales y finales han desaparecido por completo de su desarrollo, con el gametofito femenino el proceso evolutivo continúa experimentando. De ahí un número significativo de modelos del gametofito femenino y formas de su formación, en los que, sin embargo, no es posible detenerse aquí.
Literatura
1. http://ru. wikipedia.org/wiki
2. I.I. Andreeva, L.S. Rodman "Botanica" / Moscú "Coloso" 2002
3. P. Raven, R. Evert, S. Eichhorn “Botánica moderna” / Moscú “Mir” 1990
4. Mageshwari P., Embriología de angiospermas, trad. Del inglés, M., 1954; Takhtadzhyan A.L., Plantas superiores, volumen 1, M. - L., 1956;
5. Poddubnaya-Arnoldi V.A., Embriología general de angiospermas, M., 1964;
6. Savchenko M.I., Morfología del óvulo de las angiospermas, Leningrado, 1973.
Las familias Chastukhova pueden servir como buen material de reproducción. La familia Chastuha, que incluye 13-14 géneros y alrededor de 100 especies, incluye plantas tan extendidas en Rusia en las orillas de embalses y pantanos como la punta de flecha común (Sagittaria sagittifolia) con hojas en forma de flecha y racimos de flores bastante grandes, casi blancas, y el plátano. chastuha, o común (Alisma...
... (células muertas conductoras de agua con membranas lignificadas), células vivas del parénquima (realizan la transferencia de productos de la fotosíntesis desde el mesófilo y están adyacentes a la parte del floema del haz). 6.Estructura anatómica de la raíz de las monocotiledóneas La raíz, al igual que el tallo, es un órgano de simetría radial caracterizado por un crecimiento apical, realizado por actividad en su extremo del meristemo e implicado en...


La forma del brote u hojas largas (en helechos). 2. Trabajo experimental sobre la formación de actividades educativas en el proceso de estudio de la morfología de las plantas colgantes en las lecciones de biología 2.1 Métodos para estudiar las plantas colgantes en un curso escolar de biología Objetivos de la lección: formar el concepto de tallo como parte axial de un brote modificado de plantas colgantes. Revelar la relación entre...
Regenera rápidamente las capas exteriores del aura, facilitando el entendimiento mutuo. 2. Árboles de hoja caduca. Familia del abedul (Betulaceae), género Abedul (Betula). En la región de Murmansk crecen dos tipos de abedules: el abedul plateado (Betula pendula) y el abedul velloso (Betula pubescens). Con fines medicinales se utilizan yemas, hojas, corteza y alquitrán de abedul. El té de hojas de abedul es el mejor remedio para...
Óvulo u óvulo, formación multicelular en las plantas con semillas a partir de la cual se desarrolla la semilla. Las partes principales de S. son la nucela, el tegumento (o tegumentos) y el aquenio. La nucela típicamente surge en forma de tubérculo a partir de las células de la megasporofila (carpelo) de la placenta. El tegumento se forma en forma de cresta anular en la base de la nucela y crece demasiado sobre la nucela en desarrollo, dejando sobre su vértice un canal estrecho: el micropilo o pasaje de polen, debajo del cual en la mayoría de las gimnospermas hay una cámara de polen. El tallo de la semilla (funículo) se conecta. con la placenta. la parte basal desde donde se extiende el tallo de la semilla, llamada chalaza.
En el ovario del pistilo hay pequeñas formaciones: óvulos. Su número oscila entre uno (trigo, ciruela) y varios millones (en los archidae). Las funciones del óvulo son la megasporogénesis (creación de megasporas) y la megagametogénesis (formación del gametofito femenino, proceso de fecundación). El óvulo fertilizado se convierte en una semilla. La placenta es el lugar donde el óvulo se une al carpelo.
Partes del óvulo:
● nucellus, núcleo del óvulo;
● funículo, el pedúnculo con el que se une el óvulo a la placenta;
● tegumentos, cubiertas del óvulo, que forman un canal en el ápice de la nucela;
● micropilo, paso del polen;
● chalaza, la parte basal del óvulo, donde se fusionan la nucela y los tegumentos;
● hilio: el lugar donde el óvulo se une al pedúnculo.
Arroz. Estructura del óvulo
Tipos de óvulo:
ortotrópico: recto, funículo y micropilo están ubicados en los extremos opuestos del eje del óvulo (trigo sarraceno, nuez);
anatrópico (inverso): la nucela gira 180 con respecto al eje directo del óvulo, como resultado de lo cual el micropilo y el funículo se ubican cerca (angiospermas)
hemitrópico (semi-rotado): el óvulo gira 90, como resultado de lo cual el micropilo y la nucela se ubican en relación con el funículo en un ángulo de 90 (prímulas, norica).
campilotrópico (curvado unilateralmente): la nucela está curvada unilateralmente con un extremo micropilar, respectivamente, el micropilo y el funículo se encuentran cerca (leguminosas, malváceas)
anfitrópico (curvado bilateralmente): la nucela está curvada bilateralmente en forma de herradura, con el micropilo y el funículo ubicados cerca (morera, jara).

Arroz. Principales tipos de óvulos
Desarrollo del óvuloLa megasporogénesis ocurre en la esfera reproductiva femenina, en el gineceo. Morfológicamente, el gineceo está representado por un pistilo (o pistilos). La composición del pistilo incluye: estigma, estilo y ovario. El ovario contiene óvulos (uno o más). El contenido interno del óvulo es una nucela. El tegumento del óvulo está formado por un tegumento doble o único. La nucellus del óvulo contiene una célula archesporial (2n), capaz de dividirse por meiosis (en los sauces y algunas otras plantas el archesporium es multicelular). Como resultado de la meiosis, se forman cuatro megasporas haploides (n) a partir de la célula archesporial (célula madre de las megasporas). Pronto tres de ellos mueren y uno aumenta de tamaño y se divide tres veces mediante mitosis. Como resultado, se forma un saco embrionario de ocho núcleos (gametofito femenino). Tres núcleos, junto con el citoplasma adyacente, forman células antípodas, dos núcleos, un núcleo diploide central; dos núcleos - dos células sinérgicas; un núcleo se convierte en el núcleo del huevo.
Cuando un tubo polínico se acerca al óvulo, lo “siente” de antemano y se prepara para encontrarse con él. Las células compañeras comienzan a secretar sustancias mucosas. Mientras tanto, el tubo polínico crece, venciendo la resistencia de las células de las paredes del ovario. Finalmente llega al micrópilo. Se produce un proceso “dramático”: el tubo polínico perfora (y en el proceso mata) una de las células compañeras. Ambos espermatozoides abandonan el tubo polínico. El destino de la célula vegetativa de un grano de polen no es envidiable: pronto morirá. Es muy difícil observar este proceso, pero aún lo es más entender lo que sucede durante la fecundación.
En agosto de 1898, cuando aún no se conocía la doble fecundación, se celebró en Kiev el Décimo Congreso de Naturalistas y Médicos Rusos. El profesor Sergei Gavrilovich Navashin hizo un informe importante al respecto: ambos espermatozoides contenidos en el grano de polen son necesarios para el desarrollo normal de las semillas de dos especies de la familia de las liliáceas: la azucena (Lilium martagon) y el urogallo (Fritillaria tenella). ¿Por qué Navashin eligió estas plantas en particular? Probablemente porque sus espermatozoides y su saco embrionario son grandes, son fáciles de examinar bajo un microscopio. Navashev fue el primero en afirmar que el proceso de fertilización implica la fusión de dos espermatozoides con dos (!) células del saco embrionario. Pero demos la palabra al propio autor del descubrimiento.
Cada vez que se observó un tubo polínico en contacto con el saco embrionario, también se observaron ambos núcleos sexuales masculinos en el contenido del saco embrionario. Los núcleos masculinos inicialmente se encuentran uno cerca del otro.
Los núcleos masculinos luego se separan entre sí, uno penetra en el óvulo y el otro está estrechamente unido a uno de los núcleos polares que aún no se ha fusionado en este momento, es decir, el núcleo hermano del óvulo.
Mientras que el núcleo masculino está cada vez más unido al núcleo del óvulo, el núcleo polar, copulando con otro núcleo masculino, se mueve hacia otro núcleo polar, con el que se encuentra en el centro del saco embrionario.
Sólo después de la profase de división se fusionan los núcleos..."
Para aclarar la explicación, Sergei Gavrilovich hizo dibujos que gustaron mucho a los botánicos de la capital. Los llevaron para mostrarlos en una reunión de la Academia de San Petersburgo.
¿Por qué los científicos concedieron tanta importancia a este discurso? Se creía que la fertilización en las plantas ocurre de la misma manera que en los animales. Un espermatozoide y un óvulo deberían producir un cigoto a partir del cual se desarrolla una nueva planta. No es que los botánicos no hayan visto dos espermatozoides moviéndose en un tubo polínico hacia el saco embrionario. Se creía que se trataba de algún tipo de anomalía, que una planta "típica" no debería tener dos, sino un espermatozoide, y la fertilización "típica" debería ser la misma tanto en plantas como en animales. Resultó que estas ideas estaban equivocadas. La fertilización de plantas con flores es completamente diferente de la fertilización no solo de animales, sino también de otras plantas. El proceso de fertilización utilizando dos espermatozoides se llama doble fertilización. Inmediatamente después del descubrimiento de Navashin, los científicos se apresuraron a buscar medicamentos. Resultó que muchos ya habían visto la doble fertilización, pero la consideraban un proceso feo que supuestamente no producía semillas normales. Los científicos comenzaron a enviar felicitaciones a Sergei Gavrilovich. Y un científico incluso donó sus viejos preparados, que mostraban claramente una doble fertilización.
Entonces, S.G. Navashin descubrió un fenómeno asombroso. Estaba claro por qué se produce la fertilización del óvulo: para crear un cigoto y, a partir de él, una nueva planta. Pero ¿por qué fertilizar la célula central del saco embrionario? Resultó que es a partir de esta célula que se desarrolla el tejido nutritivo de la semilla, el endospermo. Navashin sugirió que el endospermo no se puede formar sin un núcleo masculino. ¿Cómo podemos probar esta suposición? Después de todo, es imposible eliminar uno de los espermatozoides del tracto polínico sin interrumpir el proceso de fertilización.
Sergei Gavrilovich decidió estudiar plantas que no tienen endospermo. ¿Qué pasa si se altera la fertilización de la célula central? Estas plantas se encuentran en la familia de las orquídeas (Orchidaceae). Las semillas de orquídeas son muy pequeñas y ni siquiera pueden germinar por sí solas (germinan solo con la ayuda de hongos basidiomicetos, formando micorrizas). Navashin vio dos espermatozoides en el tubo polínico de la orquídea. Uno de ellos fecundó el óvulo y el segundo “intentó” fertilizar la célula central con dos núcleos polares. ¡Pero en la célula central los núcleos no se fusionaron entre sí! El proceso de fertilización se vio interrumpido y, naturalmente, no se pudo formar el endospermo. En trabajos posteriores, el científico pudo demostrar que en el girasol (Helianthus annuus) y en algunas otras plantas la fertilización es doble. Después de que Navashin descubriera la doble fertilización en una variedad de plantas, concluyó que la doble fertilización es característica de todas las plantas con flores.
Echemos un vistazo a la estructura del óvulo después de la doble fertilización. Las células de ambos tegumentos tenían dos juegos de cromosomas y los retenían. Ambos conjuntos de estas células pertenecen a la planta madre. Nucellus también porta dos juegos maternos de cromosomas. En el saco embrionario, los sinérgicos murieron y las antípodas tenían cada una un juego de cromosomas. Estos conjuntos también son maternos. El cigoto, formado por la fusión de un óvulo y un espermatozoide, porta dos juegos de cromosomas: uno de la planta paterna y otro de la planta materna. Lo más interesante es que la célula central tiene tres juegos de cromosomas: uno de la planta madre y dos de la planta madre.
Después de la doble fertilización, comienzan varios procesos: el núcleo del endospermo primario se divide para formar el endospermo, el cigoto se desarrolla hasta convertirse en el embrión, los tegumentos se desarrollan hasta convertirse en la cubierta de la semilla y la pared del ovario y las estructuras asociadas forman el fruto. En las primeras etapas de desarrollo, la secuencia de divisiones celulares en embriones dicotiledóneos y monocotiledóneos es similar; en ambos casos se forman cuerpos esféricos. Posteriormente aparece una diferencia: el embrión de las dicotiledóneas tiene dos cotiledones, mientras que el de las monocotiledóneas tiene uno solo. En las semillas de algunos grupos de angiospermas, la nucela se convierte en un tejido de almacenamiento llamado peristermo. Algunas semillas contienen tanto endospermo como peristermo, como en la remolacha (Belta). Sin embargo, en muchas dicotiledóneas y algunas monocotiledóneas, todo o la mayor parte del tejido de reserva es absorbido por el embrión en desarrollo incluso antes de que la semilla entre en estado latente (en guisantes, frijoles, etc.). Los embriones de estas semillas suelen desarrollar cotiledones carnosos llenos de nutrientes. Los principales nutrientes almacenados en las semillas son los carbohidratos, las proteínas y los lípidos. Las semillas de gimnospermas y angiospermas difieren en el origen de estas sustancias. En el primero son producidos por el gametofito femenino, en el segundo por el endospermo, que no es tejido gametofítico ni esporofito.
El desarrollo del óvulo en semilla va acompañado de la transformación del ovario (y a veces de otras partes de la planta) en fruto. Durante este proceso, la pared del ovario (pericarpio o pericarpio) a menudo se espesa y se diferencia en capas separadas: el exocarpio externo (extracarpio), el mesocarpio medio (intercarpio) y el endocarpio interno (intracarpio), generalmente más visible en los carnosos que en los carnosos. en frutos secos.
Así es como se desarrolla el gametofito, la fertilización y la formación de semillas ocurren en una planta con flores "típica", pero el mundo vegetal es muy diverso y en muchas angiospermas el desarrollo se desarrolla de manera diferente, pero no menos interesante y sorprendente. Todos estos métodos de desarrollo ayudan a cada planta a mantener su estrategia en la lucha por la existencia; La ventaja de la doble fertilización no se limita al hecho de que las plantas forman reservas nutricionales en el óvulo sólo bajo la condición de fertilización (sin "desperdiciar" energía en la formación de reservas nutricionales en el óvulo, que nunca será fertilizado, como suele suceder en las gimnospermas), estas diferencias en el desarrollo, que son consecuencia de la adaptación de las plantas a las condiciones del entorno, y crean esa asombrosa variedad de formas de estructura morfológica y fisiológica que observamos en las plantas con flores.
óvulo óvulo
óvulo, óvulo (óvulo), formación multicelular de plantas con semillas, a partir de las cuales se desarrolla la semilla. Minnesota. Los embriólogos consideran a S. una estructura homóloga al megasporangio de las plantas de esporas. C consta de una nucela que contiene un megasporocito, dos o un tegumentos (cubiertas) que, cuando se cierran, forman un canal estrecho, el micropilo, a través del cual el tubo polínico penetra hasta el saco embrionario, así como un funículo (óvulos), que une S. a la placenta. La parte opuesta al micropilo se llama S. chalaza. S. de plantas con flores se forma en el ovario. S. gimnospermas están desnudas, asentadas sobre megasporofilas. En la parte chalazal de S., como resultado de la diferenciación celular, se forma una hipóstasis. para plural Las plantas con flores en S. se caracterizan por tener obturadores: secciones de tejido que crecen en forma de papilas hacia el micropilo y facilitan la penetración del tubo polínico en el saco embrionario, su crecimiento y nutrición. S. se forma en la placenta en forma de tubérculo meristemático. células del exterior capa de epidermis; cerca de su ápice aparecen una o dos células arqueesporiales (archesporia) de la capa subepidérmica, y en la base aparecen tegumentos en forma de una o dos crestas anulares. El megasporocito (célula madre de las megasporas) da lugar a megasporas, de las cuales las inferiores (chalazal), con menos frecuencia las superiores (micropilares), dan lugar a hembras. gametofito (saco embrionario en plantas con flores o endospermo primario en gimnospermas). Después de la fertilización, con el inicio del desarrollo del embrión, S. se convierte en una semilla en desarrollo. Hay 5 principales. tipos de S. según la ubicación del micropilo, funículo y eje longitudinal de la nucela: ortotrópico, o directamente (en trigo sarraceno, pimienta, aroid); anatrópico, o al revés (el tipo más común, quizás el original); hemitrópico, o semi-girado (en algunas noricáceas, en prímulas); campilotrópico, o curvado unilateral (en muchas especies de clavo, legumbres, etc.); anfitrópico, o curvado bilateralmente (en algunas especies del mismo orden que el tipo anterior). S. con una nucela poderosa, tegumentos gruesos, a veces lignificantes (el llamado crassinucellate) se considera primitivo (prevalece en las gimnospermas), con una nucellus débilmente expresada (tenuinucellate) y con un tegumento, más progresivo, que surge del primero por gradual reducción (prevalece en floración). El número de S. en el ovario de las plantas con flores varía de uno (en cereales) a 1 millón (en orquídeas).
.(Fuente: “Diccionario enciclopédico biológico”. Editor en jefe M. S. Gilyarov; Consejo editorial: A. A. Babaev, G. G. Vinberg, G. A. Zavarzin y otros - 2ª ed., corregida - M.: Sov. Encyclopedia, 1986.)
óvulo(óvulo), una formación multicelular en el ovario de las plantas con semillas, a partir de la cual se desarrolla la semilla después de la fertilización. Por lo general, consta de tegumentos (instrumentos) externos e internos. No se cierran, dejando una abertura estrecha: el conducto polínico (micrópilo). El tegumento cubre una capa cerrada multicelular: la nucela, que encierra el saco embrionario. Éste, a su vez, consta de un aparato de óvulos: tres células concentradas en el extremo más cercano al tubo polínico. Uno de ellos, con un núcleo más grande, es el óvulo (gameto femenino), los otros dos son células auxiliares o sinerigudas. En el extremo opuesto al tubo polínico se desarrollan 3 células antípodas. En el medio del saco embrionario hay una célula central. El óvulo y la célula central participan en fertilización.
.(Fuente: “Biología. Enciclopedia ilustrada moderna”. Editor jefe A. P. Gorkin; M.: Rosman, 2006.)
Sinónimos:
Vea qué es "óvulo" en otros diccionarios:
Los óvulos de las cícadas difieren en tamaño (de 5 a 6 cm de longitud en algunas especies de cícadas a 5 a 7 mm en la zamia enana) y en forma. Pero al mismo tiempo, son bastante similares en las principales características de desarrollo y estructura interna. Sentado abiertamente... ... Enciclopedia biológica
Diccionario de óvulos de sinónimos rusos. óvulo sustantivo, número de sinónimos: 2 óvulo (2) ... Diccionario de sinónimos
Lo mismo que el óvulo... Gran diccionario enciclopédico
Costura; M. Botán. embrión de semilla; óvulo. * * * óvulo es lo mismo que óvulo. * * * óvulo óvulo, lo mismo que óvulo (ver óvulo)... diccionario enciclopédico
La ubicación de los óvulos en la flor del eléboro apestoso (Helleborus foetidus) Óvulo u óvulo (lat. ... Wikipedia
Óvulo óvulo, óvulo. El órgano multicelular de las plantas con semillas, a partir del cual se desarrolla la semilla, se forma en el ovario de las angiospermas; S. consiste en nucellus
óvulo- Sinónimos: óvulo de megasporangio de plantas con semillas, rodeado por uno o dos tegumentos - tegumentos. Se forma en megasporofilas o en estróbilos (conos) femeninos de gimnospermas y en angiospermas, dentro del ovario del pistilo de una flor (ver ... Anatomía y morfología de las plantas.
Lo mismo que el óvulo... Ciencias Naturales. diccionario enciclopédico
óvulo- semilla atok, tejido... diccionario de ortografía ruso
óvulo- óvulo (óvulo), una formación multicelular en los órganos reproductivos de las células seminales, a partir de la cual se desarrolla la semilla durante el desarrollo (generalmente después de la fertilización). En las angiospermas, S. se forma oculta dentro del ovario, en las gimnospermas se ubica... ... Diccionario enciclopédico agrícola
- En contacto con 0
- Google+ 0
- DE ACUERDO 0
- Facebook 0






