Kahe neuroni kokkupuuteala nimetatakse sünaps.
Aksodendriitse sünapsi sisemine struktuur.A) Elektrilised sünapsid. Elektrilised sünapsid on imetajate närvisüsteemis haruldased. Need moodustuvad külgnevate neuronite dendriitide või somaatiliste vahekohtade (nexuse) kaudu, mis on ühendatud tsütoplasmaatiliste kanalitega, mille läbimõõt on 1,5 nm. Signaali edastamise protsess toimub ilma sünaptilise viivituseta ja ilma vahendajate osaluseta.
Elektriliste sünapside kaudu võivad elektrotoonilised potentsiaalid levida ühelt neuronilt teisele. Tiheda sünaptilise kontakti tõttu on signaali edastamise moduleerimine võimatu. Nende sünapside ülesanne on samaaegselt ergutada neuroneid, mis täidavad sama funktsiooni. Näitena võib tuua pikliku medulla hingamiskeskuse neuronid, mis tekitavad sissehingamisel sünkroonselt impulsse. Lisaks on näiteks sakaade kontrollivad närviahelad, mille puhul pilgu fikseerimise punkt liigub ühelt tähelepanuobjektilt teisele.
b) Keemilised sünapsid. Enamik närvisüsteemi sünapse on keemilised. Selliste sünapside toimimine sõltub saatjate vabanemisest. Klassikalist keemilist sünapsi esindavad presünaptiline membraan, sünaptiline lõhe ja postsünaptiline membraan. Presünaptiline membraan on signaali edastava raku närvilõpme klubikujulise laiendi osa, postsünaptiline membraan aga signaali vastuvõtva raku osa.
Saatja vabaneb klavaadi pikendusest eksotsütoosi teel, läbib sünaptilise lõhe ja seondub postsünaptilise membraani retseptoritega. Postsünaptilise membraani all on subsünaptiline aktiivne tsoon, milles pärast postsünaptilise membraani retseptorite aktiveerimist toimuvad mitmesugused biokeemilised protsessid.
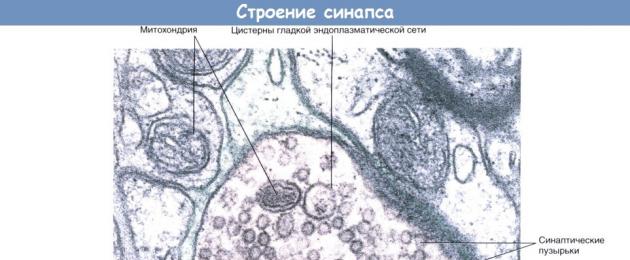
Klubikujuline pikendus sisaldab mediaatoreid sisaldavaid sünaptilisi vesiikuleid, aga ka suurt hulka sileda endoplasmaatilise retikulumi mitokondreid ja tsisterneid. Traditsiooniliste fikseerimistehnikate kasutamine rakkude uurimisel võimaldab eristada presünaptilisi tihendeid presünaptilisel membraanil, piirates sünapsi aktiivseid tsoone, kuhu sünaptilised vesiikulid mikrotuubulite abil suunatakse.
 Aksodendriitne sünaps.
Aksodendriitne sünaps. Seljaaju proovi osa: sünaps dendriidi terminaalse osa ja arvatavasti motoorse neuroni vahel.
Eksitatoorsetele sünapsidele on iseloomulik ümarate sünaptiliste vesiikulite olemasolu ja postsünaptiline tihendus.
Dendriit lõigati põikisuunas, mida tõendab paljude mikrotuubulite olemasolu.
Lisaks on näha mõningaid neurofilamente. Sünapsi asukohta ümbritseb protoplasmaatiline astrotsüüt.
 Protsessid, mis toimuvad kahte tüüpi närvilõpmetes.
Protsessid, mis toimuvad kahte tüüpi närvilõpmetes. (A) Väikeste molekulide (nt glutamaat) sünaptiline ülekanne.
(1) Sünaptiliste vesiikulite membraanvalke sisaldavad transportvesiikulid suunatakse mööda mikrotuubuleid nuiakujulise paksenemise plasmamembraanile.
Samal ajal viiakse ensüümi ja glutamaadi molekulid üle aeglase transpordi teel.
(2) Vesiikulite membraani valgud väljuvad plasmamembraanist ja moodustavad sünaptilisi vesiikuleid.
(3) Glutamaat laetakse sünaptilistesse vesiikulitesse; toimub vahendaja kuhjumine.
(4) Glutamaati sisaldavad vesiikulid lähenevad presünaptilisele membraanile.
(5) Depolarisatsiooni tulemusena toimub osaliselt hävinud vesiikulitest vahendaja eksotsütoos.
(6) Vabanenud saatja levib difuusselt sünaptilise lõhe piirkonnas ja aktiveerib spetsiifilisi retseptoreid postsünaptilisel membraanil.
(7) Sünaptiliste vesiikulite membraanid transporditakse endotsütoosi teel tagasi rakku.
(8) Glutamaadi osaline tagasihaaramine rakku toimub taaskasutamiseks.
(B) Neuropeptiidide (nt aine P) ülekanne, mis toimub samaaegselt sünaptilise ülekandega (nt glutamaat).
Nende ainete ühine ülekanne toimub unipolaarsete neuronite kesknärvilõpmetes, mis tagavad valutundlikkuse.
(1) Golgi kompleksis (perikarüoni piirkonnas) sünteesitud vesiikulid ja peptiidide prekursorid (propeptiidid) transporditakse kiirtranspordiga klubikujulisele pikendusele.
(2) Kui nad sisenevad klubikujulise paksenemise piirkonda, on peptiidimolekuli moodustumise protsess lõppenud ja vesiikulid transporditakse plasmamembraanile.
(3) Membraani depolarisatsioon ja vesiikulite sisu viimine rakkudevahelisse ruumi eksotsütoosi teel.
(4) Samal ajal vabaneb glutamaat.
1. Retseptori aktiveerimine. Transmittermolekulid läbivad sünaptilist pilu ja aktiveerivad postsünaptilisel membraanil paarikaupa paiknevad retseptorvalgud. Retseptorite aktiveerimine käivitab ioonsed protsessid, mis põhjustavad postsünaptilise membraani depolarisatsiooni (ergastav postsünaptiline toime) või postsünaptilise membraani hüperpolarisatsiooni (inhibeeriv postsünaptiline toime). Elektrotoonilisuse muutus kandub sooma edasi elektrotoonilise potentsiaalina, mis levides laguneb, mille tõttu muutub puhkepotentsiaal aksoni algsegmendis.
Ioonprotsesse kirjeldatakse üksikasjalikult veebisaidi eraldi artiklis. Kui domineerivad ergastavad postsünaptilised potentsiaalid, depolariseeritakse aksoni esialgne segment lävitasemeni ja tekitab aktsioonipotentsiaali.
Kesknärvisüsteemi kõige levinum ergastav neurotransmitter on glutamaat ja inhibeeriv gamma-aminovõihape (GABA). Perifeerses närvisüsteemis toimib atsetüülkoliin vöötlihaste motoorsete neuronite ja glutamaat sensoorsete neuronite edastajana.
Glutamatergilistes sünapsides toimuvate protsesside jada on näidatud alloleval joonisel. Kui glutamaat viiakse koos teiste peptiididega, toimub peptiidide vabanemine ekstrasünaptiliste radade kaudu.
Enamik sensoorseid neuroneid eritavad lisaks glutamaadile ka teisi peptiide (üht või mitut), mis vabanevad neuroni erinevates osades; nende peptiidide põhiülesanne on aga moduleerida (suurendada või vähendada) sünaptilise glutamaadi ülekande efektiivsust.
Lisaks võib neurotransmissioon toimuda difuusse ekstrasünaptilise signaaliülekande kaudu, mis on iseloomulik monoaminergilistele neuronitele (neuronid, mis kasutavad neurotransmissiooni vahendamiseks biogeenseid amiine). On kahte tüüpi monoamiinergilised neuronid. Mõnedes neuronites sünteesitakse aminohappest türosiinist katehhoolamiinid (norepinefriin või dopamiin), teistes aga serotoniin aminohappest trüptofaanist. Näiteks dopamiin vabaneb nii sünaptilises piirkonnas kui ka aksonite veenilaienditest, mille puhul toimub ka selle neurotransmitteri süntees.
Dopamiin tungib kesknärvisüsteemi rakkudevahelisse vedelikku ja on enne lagunemist võimeline aktiveerima spetsiifilisi retseptoreid kuni 100 mikroni kaugusel. Monoaminergilised neuronid esinevad paljudes kesknärvisüsteemi struktuurides; nende neuronite impulsi ülekande katkemine põhjustab mitmesuguseid haigusi, sealhulgas Parkinsoni tõbe, skisofreeniat ja suurt depressiooni.
Lämmastikoksiid (gaasiline molekul) osaleb ka difuusses neurotransmissioonis glutamatergilises neuronaalses süsteemis. Liigsel lämmastikoksiidil on tsütotoksiline toime, eriti neis piirkondades, kus verevarustus on arteriaalse tromboosi tõttu häiritud. Glutamaat on ka potentsiaalselt tsütotoksiline neurotransmitter.
Erinevalt difuussest neurotransmissioonist nimetatakse traditsioonilist sünaptilist signaaliülekannet selle suhtelise stabiilsuse tõttu "juhiks".
V) Kokkuvõte. Kesknärvisüsteemi multipolaarsed neuronid koosnevad somast, dendriitidest ja aksonist; akson moodustab tagatis- ja terminaliharud. Soma sisaldab sileda ja karedat endoplasmaatilist retikulumit, Golgi komplekse, neurofilamente ja mikrotuubuleid. Mikrotuubulid tungivad läbi kogu neuroni, osalevad sünaptiliste vesiikulite, mitokondrite ja membraani moodustavate ainete anterograadse transpordi protsessis ning tagavad ka "marker"-molekulide ja hävitatud organellide retrograadse transpordi.
On olemas kolme tüüpi keemilisi interneuronaalseid interaktsioone: sünaptiline (nt glutamatergiline), ekstrasünaptiline (peptidergiline) ja difuusne (nt monoaminergiline, serotonergiline).
Keemilised sünapsid liigitatakse nende anatoomilise struktuuri järgi aksodendriitilisteks, aksosomaatilisteks, aksoaksonaalseteks ja dendrodendriitideks. Sünapsi esindavad pre- ja postsünaptilised membraanid, sünaptiline lõhe ja subsünaptiline aktiivne tsoon.
Elektrilised sünapsid tagavad tervete rühmade üheaegse aktiveerimise, moodustades nende vahel elektriühendusi lünklike kontaktide (nexuse) tõttu.
Glutamatergiliste (1) ja dopamiinergiliste (2) neuronite aksonid moodustavad tihedad sünaptilised kontaktid juttkeha stellaatneuroni (3) protsessiga.
Dopamiin ei eraldu mitte ainult presünaptilisest piirkonnast, vaid ka aksoni varikoossest paksenemisest, kust see difundeerub rakkudevahelisse ruumi ja aktiveerib dendriittüve ja kapillaaride peritsüütide seinte dopamiini retseptoreid.
 Deinhibeerimine.
Deinhibeerimine. (A) Eksitatoorne neuron 1 aktiveerib inhibeeriva neuroni 2, mis omakorda pärsib neuronit 3.
(B) Teise inhibeeriva neuroni (2b) ilmumisel on neuronile 3 vastupidine mõju, kuna neuron 2b on inhibeeritud.
Spontaanselt aktiivne neuron 3 genereerib signaale inhibeerivate mõjude puudumisel.
2. Ravimid – “võtmed” ja “lukud”. Retseptorit saab võrrelda lukuga ja vahendajat sellele vastava võtmega. Kui vahendaja vabanemise protsess on vanuse või mõne haiguse tagajärjel häiritud, võib ravim mängida "varuvõtme" rolli, mis täidab vahendajaga sarnast funktsiooni. Seda ravimit nimetatakse agonistiks. Samal ajal võib ülemäärase tootmise korral vahendaja "peatada" retseptori blokaatoriga - "võltsvõtmega", mis võtab ühendust "luku" retseptoriga, kuid ei põhjusta selle aktiveerimist.
3. Pidurdamine ja deaktiveerimine. Spontaanselt aktiivsete neuronite toimimist pärsib inhibeerivate neuronite (tavaliselt GABAergiliste) mõju. Inhibeerivate neuronite aktiivsust võivad omakorda pärssida teised neile mõjuvad inhibeerivad neuronid, mille tulemuseks on sihtraku inhibeerimine. Desinhibeerimisprotsess on basaalganglionide neuronaalse aktiivsuse oluline tunnus.
4. Haruldased keemilised sünapsi tüübid. Aksoaksonaalseid sünapse on kahte tüüpi. Mõlemal juhul moodustab klubikujuline paksene inhibeeriva neuroni. Esimest tüüpi sünapsid moodustuvad aksoni algsegmendi piirkonnas ja edastavad inhibeeriva neuroni võimsa inhibeeriva toime. Teist tüüpi sünapsid moodustuvad inhibeeriva neuroni klubikujulise paksenemise ja ergastavate neuronite klubikujulise paksenemise vahel, mis viib saatjate vabanemise pärssimiseni. Seda protsessi nimetatakse presünaptiliseks inhibeerimiseks. Sellega seoses pakub traditsiooniline sünaps postsünaptilist inhibeerimist.
Dendro-dendriitsed (D-D) sünapsid moodustuvad külgnevate ogaliste neuronite dendriitide dendriitide vahel. Nende ülesanne ei ole tekitada närviimpulssi, vaid muuta sihtraku elektrilist tooni. Järjestikustes D-D sünapsides paiknevad sünaptilised vesiikulid ainult ühes dendriitses selgroos ja vastastikustes D-D sünapsides mõlemas. Ergastavad D-D sünapsid on näidatud alloleval joonisel. Inhibeerivad D-D sünapsid on laialdaselt esindatud talamuse lülitustuumades.
Lisaks on mõned somato-dendriit- ja somato-somaatilised sünapsid.
 Ajukoore aksoaksonaalsed sünapsid.
Ajukoore aksoaksonaalsed sünapsid. Nooled näitavad impulsside suunda.
 (1) ajju liikuva seljaaju neuroni presünaptiline ja (2) postsünaptiline pärssimine.
(1) ajju liikuva seljaaju neuroni presünaptiline ja (2) postsünaptiline pärssimine. Nooled näitavad impulsi juhtivuse suunda (lülitusneuroni pärssimine pärssivate mõjude mõjul on võimalik).
 Eksitatoorsed dendro-dendriitsed sünapsid. Kujutatud on kolme neuroni dendriite.
Eksitatoorsed dendro-dendriitsed sünapsid. Kujutatud on kolme neuroni dendriite. Vastastikune sünaps (paremal). Nooled näitavad elektrotooniliste lainete levimise suunda.
Õppevideo - sünapsi struktuur
Sünaps on neuronite vahelise funktsionaalse, mitte füüsilise kontakti koht; see edastab teavet ühest rakust teise. Tavaliselt on ühe neuroni aksoni terminaalsete harude ja dendriitide vahel sünapsid ( aksodendriitne sünapsid) või keha ( aksosomaatiline teise neuroni sünapsid. Sünapside arv on tavaliselt väga suur, mis annab suure ala info edastamiseks. Näiteks seljaaju üksikute motoorsete neuronite dendriitidel ja rakukehadel on üle 1000 sünapsi. Mõnel ajurakul võib olla kuni 10 000 sünapsi (joonis 16.8).
Sünapse on kahte tüüpi - elektriline Ja keemiline- sõltuvalt neid läbivate signaalide iseloomust. Motoorse neuroni klemmide ja lihaskiu pinna vahel on neuromuskulaarne ristmik, mis erinevad oma struktuurilt interneuronite sünapsidest, kuid sarnanevad nendega funktsionaalselt. Struktuurseid ja füsioloogilisi erinevusi normaalse sünapsi ja neuromuskulaarse ristmiku vahel kirjeldatakse veidi hiljem.
Keemilise sünapsi struktuur
Keemilised sünapsid on selgroogsetel kõige levinum sünapsi tüüp. Need on närvilõpmete sibulakujulised paksenemised, mida nimetatakse sünaptilised naastud ja asub dendriidi otsa vahetus läheduses. Sünaptilise naastu tsütoplasma sisaldab mitokondreid, sileda endoplasmaatilist retikulumit, mikrofilamente ja palju sünaptilised vesiikulid. Iga vesiikul on umbes 50 nm läbimõõduga ja sisaldab vahendaja- aine, millega närvisignaal edastatakse sünapsi kaudu. Sünaptilise naastu membraan sünapsi enda piirkonnas pakseneb tsütoplasma tihendamise tulemusena ja moodustub presünaptiline membraan. Dendriitmembraan sünapsi piirkonnas on samuti paksenenud ja moodustub postsünaptiline membraan. Need membraanid on eraldatud vahega - sünaptiline lõhe umbes 20 nm lai. Presünaptiline membraan on konstrueeritud nii, et selle külge saavad kinnituda sünaptilised vesiikulid ja sünaptilisse pilusse pääsevad vahendajad. Postsünaptiline membraan sisaldab suuri valgumolekule, mis toimivad retseptorid vahendajad ja paljud kanalid Ja poorid(tavaliselt suletud), mille kaudu võivad ioonid siseneda postsünaptilisse neuronisse (vt joon. 16.10, A).
Sünaptilised vesiikulid sisaldavad saatjat, mis moodustub kas neuroni kehas (ja siseneb sünaptilisse naastu, läbides kogu aksoni) või otse sünaptilisse naastu. Mõlemal juhul on vahendaja sünteesiks vaja ensüüme, mis moodustuvad rakukehas ribosoomidel. Sünaptilises naastudes pakitakse saatjamolekulid vesiikulitesse, milles neid hoitakse kuni vabanemiseni. Selgroogsete närvisüsteemi peamised vahendajad on atsetüülkoliin Ja norepinefriin, kuid on ka teisi vahendajaid, millest tuleb juttu hiljem.
Atsetüülkoliin on ammooniumi derivaat, mille valem on näidatud joonisel fig. 16.9. See on esimene teadaolev vahendaja; 1920. aastal eraldas Otto Lewy selle konna südame vagusnärvi parasümpaatiliste neuronite otstest (jaotis 16.2). Norepinefriini struktuuri käsitletakse üksikasjalikult jaotises. 16.6.6. Neuroneid, mis vabastavad atsetüülkoliini, nimetatakse kolinergiline ja need, mis vabastavad norepinefriini - adrenergiline.

Sünaptilise ülekande mehhanismid
Arvatakse, et närviimpulsi saabumine sünaptilisele naastule põhjustab presünaptilise membraani depolarisatsiooni ja selle Ca 2+ ioonide läbilaskvuse suurenemise. Sünaptilisse naastu sisenevad Ca 2+ ioonid põhjustavad sünaptiliste vesiikulite liitumist presünaptilise membraaniga ja nende sisu vabanemist rakust (eksotsütoos), mille tulemusena satub sünaptilisse pilusse. Kogu seda protsessi nimetatakse elektrosekretoorne sidestus. Kui vahendaja vabaneb, kasutatakse vesiikulite materjali uute vesiikulite moodustamiseks, mis on täidetud vahendaja molekulidega. Iga viaal sisaldab umbes 3000 atsetüülkoliini molekuli.
Vahendajamolekulid difundeeruvad läbi sünaptilise pilu (see protsess võtab aega umbes 0,5 ms) ja seonduvad postsünaptilisel membraanil paiknevate retseptoritega, mis on võimelised ära tundma atsetüülkoliini molekulaarstruktuuri. Kui retseptormolekul seostub saatjaga, muutub selle konfiguratsioon, mis viib ioonikanalite avanemiseni ja ioonide sisenemiseni postsünaptilisse rakku, põhjustades depolarisatsioon või hüperpolarisatsioon(joon. 16.4, A) selle membraan, olenevalt vabanenud vahendaja olemusest ja retseptori molekuli struktuurist. Transmittermolekulid, mis põhjustavad muutust postsünaptilise membraani läbilaskvuses, eemaldatakse kohe sünaptilisest pilust kas reabsorptsiooni teel presünaptilise membraani poolt või difusiooni teel pilust või ensümaatilise hüdrolüüsi teel. Millal kolinergiline sünapsides hüdrolüüsitakse ensüümi toimel sünaptilises pilus paiknev atsetüülkoliin atsetüülkoliinesteraas, lokaliseeritud postsünaptilisel membraanil. Hüdrolüüsi tulemusena tekib koliin, mis imendub tagasi sünaptilisse naastu ja muundub seal uuesti atsetüülkoliiniks, mis ladestub vesiikulites (joon. 16.10).

IN stimuleeriv Sünapsides avanevad atsetüülkoliini mõjul spetsiifilised naatriumi- ja kaaliumikanalid ning Na + ioonid sisenevad rakku ja K + ioonid lahkuvad sellest vastavalt nende kontsentratsioonigradientidele. Selle tulemusena toimub postsünaptilise membraani depolarisatsioon. Seda depolarisatsiooni nimetatakse ergastav postsünaptiline potentsiaal(EPSP). EPSP amplituud on tavaliselt väike, kuid selle kestus on pikem kui aktsioonipotentsiaalil. EPSP amplituud muutub järk-järgult, mis viitab sellele, et saatja vabaneb osadena või "kvantidena", mitte üksikute molekulide kujul. Ilmselt vastab iga kvant saatja vabanemisele ühest sünaptilisest vesiikulist. Üks EPSP ei ole reeglina võimeline tekitama aktsioonipotentsiaali tekkimiseks vajaliku läviväärtuse depolarisatsiooni. Kuid mitmete EPSP-de depolariseerivad mõjud liidetakse kokku ja seda nähtust nimetatakse summeerimine. Kaks või enam EPSP-d, mis esinevad samaaegselt sama neuroni erinevates sünapsides, võivad ühiselt tekitada depolarisatsiooni, mis on piisav postsünaptilise neuroni aktsioonipotentsiaali ergutamiseks. Seda nimetatakse ruumiline summeerimine. Transmitteri kiirelt korduv vabanemine sama sünaptilise naastu vesiikulitest intensiivse stiimuli mõjul põhjustab üksikuid EPSP-sid, mis ajas nii sageli üksteisele järgnevad, et ka nende mõjud summeeritakse ja tekitavad postsünaptilises neuronis aktsioonipotentsiaali. Seda nimetatakse aja summeerimine. Seega võivad impulsid tekkida ühes postsünaptilises neuronis kas mitme seotud presünaptilise neuroni nõrga stimulatsiooni tulemusena või ühe selle presünaptilise neuroni korduva stimulatsiooni tulemusena. IN pidur sünapsides suurendab saatja vabanemine postsünaptilise membraani läbilaskvust tänu spetsiifiliste kanalite avanemisele K + ja Cl - ioonide jaoks. Kontsentratsioonigradiente mööda liikudes põhjustavad need ioonid membraani hüperpolarisatsiooni, nn inhibeeriv postsünaptiline potentsiaal(TPSP).
Vahendajatel endil ei ole ergastavaid ega inhibeerivaid omadusi. Näiteks atsetüülkoliinil on ergastav toime enamikes neuromuskulaarsetes ühenduskohtades ja muudes sünapsides, kuid see põhjustab pärssimist südame ja vistseraalsete lihaste neuromuskulaarsetes ühenduskohtades. Need vastandlikud mõjud on tingitud sündmustest, mis ilmnevad postsünaptilisel membraanil. Retseptori molekulaarsed omadused määravad, millised ioonid postsünaptilisse neuronisse sisenevad, ja need ioonid omakorda määravad postsünaptiliste potentsiaalide muutuse olemuse, nagu eespool kirjeldatud.
Elektrilised sünapsid
Paljudel loomadel, sealhulgas koelenteraatidel ja selgroogsetel, toimub impulsside ülekandmine mõne sünapsi kaudu elektrivoolu läbimise teel pre- ja postsünaptiliste neuronite vahel. Nende neuronite vahelise pilu laius on vaid 2 nm ning kogutakistus membraanide voolule ja tühimikku täitvale vedelikule on väga väike. Impulsid läbivad sünapsid viivitamatult ning nende ülekandumist ei mõjuta ravimid ega muud kemikaalid.
Neuromuskulaarne ristmik
Neuromuskulaarne ristmik on spetsiifiline sünapsi tüüp motoorse neuroni (motoneuroni) otste ja endomüsium lihaskiud (jaotis 17.4.2). Igal lihaskiul on spetsiaalne ala - mootori otsaplaat, kus motoorse neuroni (motoneuroni) akson hargneb, moodustades umbes 100 nm paksuseid müeliniseerimata oksi, mis kulgevad madalates soontes piki lihasmembraani pinda. Lihasraku membraan – sarkolemma – moodustab palju sügavaid volte, mida nimetatakse postsünaptiliseks voldiks (joon. 16.11). Motoorsete neuronite terminalide tsütoplasma on sarnane sünaptilise naastu sisuga ja vabastab stimulatsiooni ajal atsetüülkoliini, kasutades sama mehhanismi, mida käsitleti eespool. Sarkolemma pinnal paiknevate retseptormolekulide konfiguratsiooni muutused toovad kaasa muutuse selle Na + ja K + läbilaskvuses ning selle tulemusena toimub lokaalne depolarisatsioon, nn. otsaplaadi potentsiaal(PKP). See depolarisatsioon on piisavalt suur, et tekitada aktsioonipotentsiaal, mis levib mööda sarkolemma sügavale kiudu mööda põikisuunaliste tuubulite süsteemi ( T-süsteem) (jaotis 17.4.7) ja põhjustab lihaste kokkutõmbumist.

Sünapside ja neuromuskulaarsete ühenduste funktsioonid
Interneuronite sünapside ja neuromuskulaarsete ühenduste põhiülesanne on signaalide edastamine retseptoritelt efektoritele. Lisaks määravad nende keemilise sekretsiooni kohtade struktuur ja korraldus mitmed olulised närviimpulsside juhtimise tunnused, mille võib kokku võtta järgmiselt:
1. Ühesuunaline ülekanne. Saatja vabanemine presünaptilisest membraanist ja retseptorite lokaliseerimine postsünaptilisel membraanil võimaldavad edastada närvisignaale mööda seda teed ainult ühes suunas, mis tagab närvisüsteemi töökindluse.
2. Kasu. Iga närviimpulss põhjustab piisava atsetüülkoliini vabanemise neuromuskulaarses ristmikul, et tekitada lihaskius leviv reaktsioon. Tänu sellele võivad neuromuskulaarsesse liitumiskohta saabuvad närviimpulsid, ükskõik kui nõrgad, tekitada efektorreaktsiooni ja see tõstab süsteemi tundlikkust.
3. Kohanemine või majutus. Pideva stimulatsiooni korral väheneb sünapsis vabaneva saatja hulk järk-järgult, kuni saatja reservid on ammendatud; siis nad ütlevad, et sünaps on väsinud ja signaalide edasine edastamine sellele on pärsitud. Väsimuse adaptiivne väärtus seisneb selles, et see hoiab ära efektori kahjustamise üleergutuse tõttu. Kohanemine toimub ka retseptori tasandil. (Vt kirjeldust jaotises 16.4.2.)
4. Integratsioon. Postsünaptiline neuron võib vastu võtta signaale suurelt hulgalt ergastavatelt ja inhibeerivatelt presünaptilistest neuronitest (sünaptiline konvergents); sel juhul on postsünaptiline neuron võimeline kõigi presünaptiliste neuronite signaalid kokku võtma. Ruumilise summeerimise kaudu integreerib neuron signaale paljudest allikatest ja annab koordineeritud vastuse. Mõne sünapsi puhul toimub hõlbustamine, mille korral sünaps muutub pärast iga stiimulit tundlikumaks järgmise stiimuli suhtes. Seetõttu võivad järjestikused nõrgad stiimulid esile kutsuda vastuse ja seda nähtust kasutatakse teatud sünapside tundlikkuse suurendamiseks. Hõlbustamist ei saa pidada ajutiseks liitmiseks: postsünaptilises membraanis toimub keemiline muutus, mitte postsünaptilise membraani potentsiaalide elektriline liitmine.
5. Diskrimineerimine. Ajaline summeerimine sünapsis võimaldab nõrgad taustaimpulsid välja filtreerida enne, kui need jõuavad ajju. Näiteks naha, silmade ja kõrvade eksteroretseptorid saavad pidevalt keskkonnast signaale, mis ei ole närvisüsteemi jaoks eriti olulised: ainult tema jaoks on olulised. muudatusi stiimuli intensiivsus, mis viib impulsside sageduse suurenemiseni, mis tagab nende edasikandumise sünapsi kaudu ja sobiva reaktsiooni.
6. Pidurdamine. Teatud postsünaptilisele membraanile mõjuvad blokeerivad ained võivad pärssida signaaliülekannet sünapside ja neuromuskulaarsete ühenduste kaudu (vt allpool). Presünaptiline inhibeerimine on võimalik ka siis, kui aksoni lõpus, vahetult antud sünapsi kohal, lõpeb teine akson, moodustades siin inhibeeriva sünapsi. Sellise inhibeeriva sünapsi stimuleerimisel väheneb esimeses ergastavas sünapsis väljutatavate sünaptiliste vesiikulite arv. Selline seade võimaldab muuta antud presünaptilise neuroni mõju, kasutades teisest neuronist tulevaid signaale.
Keemiline mõju sünapsile ja neuromuskulaarsele ristmikule
Kemikaalid täidavad närvisüsteemis palju erinevaid funktsioone. Mõnede ainete mõju on laialt levinud ja hästi uuritud (näiteks atsetüülkoliini ja adrenaliini stimuleeriv toime), teiste mõju on aga lokaalne ega ole veel hästi mõistetav. Mõned ained ja nende funktsioonid on toodud tabelis. 16.2.

Arvatakse, et mõned psüühikahäirete, näiteks ärevuse ja depressiooni raviks kasutatavad ravimid mõjutavad keemilist ülekannet sünapsides. Paljud rahustid ja rahustid (tritsüklilised antidepressandid imipramiin, reserpiin, monoamiini oksüdaasi inhibiitorid jne) avaldavad oma ravitoimet interakteerudes mediaatorite, nende retseptorite või üksikute ensüümidega. Näiteks monoamiini oksüdaasi inhibiitorid inhibeerivad adrenaliini ja norepinefriini lagundamisel osalevat ensüümi ning avaldavad suure tõenäosusega oma terapeutilist toimet depressioonile, suurendades nende vahendajate toime kestust. Hallutsinogeenide tüüp Lüsergiinhappe dietüülamiid Ja meskaliin, reprodutseerida mõnede looduslike aju vahendajate toimet või pärssida teiste vahendajate toimet.
Hiljutised uuringud teatud valuvaigistite, mida nimetatakse opiaatideks, mõju kohta heroiini Ja morfiin- näitas, et imetajate aju sisaldab looduslikku (endogeenne) ained, mis põhjustavad sarnast toimet. Kõiki neid aineid, mis interakteeruvad opiaadiretseptoritega, nimetatakse ühiselt endorfiinid. Praeguseks on avastatud palju selliseid ühendeid; Neist kõige paremini uuritud suhteliselt väikeste peptiidide rühm nn enkefaliinid(met-enkefaliin, β-endorfiin jne). Arvatakse, et need suruvad valu alla, mõjutavad emotsioone ja on seotud mõne vaimuhaigusega.
Kõik see on avanud uusi võimalusi aju funktsioonide ja valule avalduva mõju aluseks olevate biokeemiliste mehhanismide uurimiseks ja ravimiseks, kasutades selliseid erinevaid meetodeid nagu sugestioon, hüpno? ja nõelravi. Paljud teised ained, nagu endorfiinid, tuleb veel eraldada ning nende struktuur ja funktsioonid kindlaks määrata. Nende abiga on võimalik saada täielikum arusaam aju toimimisest ja see on vaid aja küsimus, kuna nii väikestes kogustes esinevate ainete eraldamise ja analüüsimise meetodeid täiustatakse pidevalt.
Loeng 2. Sünapside füsioloogia: struktuur, klassifikatsioon ja toimemehhanismid. Vahendajad, käitumise neurokeemilised alused.
19. sajandi lõpus eksisteeris kaks paralleelset närvisüsteemi (NS) organisatsiooni teooriat. Retikulaarne teooria arvati, et NS on funktsionaalne süntsüüt: neuronid on ühendatud protsesside kaudu, mis on sarnased vereringesüsteemi kapillaaridega. Vastavalt Waldeyeri rakuteooria(1981) NS koosneb üksikutest membraanidega eraldatud neuronitest. Üksikute neuronite vahelise interaktsiooni probleemi lahendamiseks Sherrington 1987. aastal soovitas ta spetsiaalse membraani moodustumise olemasolu - sünaps. Elektronmikroskoobi abil kinnitati sünapside olemasolu ühemõtteliselt. NS struktuuri rakuteooria sai aga üldtunnustatud; iroonilisel kombel avastasid Fershpan ja Potter 1959. aastal koorikloomade NS-s vaheühendustega sünapsi (elektrilise sünapsi).
Sünaps on kahest (või enamast) rakust koosnev membraanmoodustis, milles ergastus (informatsioon) kandub ühest rakust teise.
Sünapside klassifikatsioon on järgmine:
1) ergastuse ülekandemehhanismi (ja struktuuri järgi):
Keemiline;
Elektriseadmed (ephaps);
Segatud.
2) vastavalt vabanenud neurotransmitterile:
Adrenergiline – neurotransmitter norepinefriin;
Koliinergiline – neurotransmitter atsetüülkoliin;
Dopamiinergiline – neurotransmitter dopamiin;
Serotonergiline – neurotransmitter serotoniin;
GABAergic – neurotransmitter gamma-aminovõihape (GABA)
3) mõjul:
Põnev;
Pidur.
4) asukoha järgi:
Neuromuskulaarne;
Neuro-närvi:
a) aksosomaatiline;
b) aksoaksonaalne;
c) aksodendriitne;
d) dendrosomaatiline.
Vaatleme kolme tüüpi sünapsi: keemiline, elektriline ja segatud(ühendades keemiliste ja elektriliste sünapside omadused).
Sõltumata tüübist on sünapsidel ühised struktuurilised tunnused: närviprotsess lõpus moodustab laiendi ( sünaptiline tahvel, SB); SB terminaalne membraan erineb teistest neuronimembraani osadest ja seda nimetatakse presünaptiline membraan(PreSM); teise raku spetsiaalset membraani nimetatakse postsünaptiliseks membraaniks (PostSM); asub sünapsi membraanide vahel sünaptiline lõhe(SCH, joon. 1, 2).
Riis. 1. Keemilise sünapsi ehituse skeem
Elektrilised sünapsid(ephapses, ES) leidub tänapäeval mitte ainult vähilaadsete, vaid ka molluskite, lülijalgsete ja imetajate NS-s. ES-il on mitmeid ainulaadseid omadusi. Neil on kitsas sünaptiline lõhe (umbes 2-4 nm), mille tõttu saab ergastust elektrokeemiliselt (nagu EMF-i tõttu närvikiu kaudu) edasi anda. suurel kiirusel ja mõlemas suunas: nii PreSM-membraanilt PostSM-ile kui ka PostSM-ilt PreSM-ile. Rakkude vahel on vaheühendused (konnexid või konneksonid), mille moodustavad kaks konneksiini valku. Iga konneksiini kuus subühikut moodustavad PreSM- ja PostSM-kanalid, mille kaudu saavad rakud vahetada madalmolekulaarseid aineid molekulmassiga 1000-2000 daltonit. Konneksonite tööd saab reguleerida Ca 2+ ioonidega (joonis 2).

Riis. 2. Elektrilise sünapsi skeem
ES on suurem spetsialiseerumine võrreldes keemiliste sünapsidega ja tagavad suure ergastuse edastuskiiruse. Siiski näib, et see on ilma jäetud võimalusest edastatud teavet peenemalt analüüsida (reguleerida).
NS-is domineerivad keemilised sünapsid. Nende uurimistöö ajalugu algab Claude Bernardi töödega, kes avaldas 1850. aastal artikli “Research on Curare”. Ta kirjutas nii: "Curare on tugev mürk, mille on valmistanud mõned Amazonase metsades elavad rahvad (peamiselt kannibalid). Ja veel: "Curare sarnaneb maomürgiga selle poolest, et seda saab karistamatult viia inimeste või loomade seedetrakti, samas kui naha alla või mis tahes kehaosasse süstimine põhjustab kiiresti surma. ...mõne hetke pärast heidavad loomad nagu väsinud pikali. Siis hingamine lakkab ning nende tundlikkus ja elu kaovad, ilma et loomad hüüaksid või valu märke näitaksid. Kuigi C. Bernard ei jõudnud närviimpulsside keemilise ülekande ideeni, võimaldasid tema klassikalised eksperimendid curare'ga sellel ideel tekkida. Möödus üle poole sajandi, kui J. Langley tuvastas (1906), et curare halvav toime on seotud lihase erilise osaga, mida ta nimetas vastuvõtlikuks aineks. Esimese ettepaneku ergastuse ülekandmise kohta närvilt efektororganile, kasutades keemilist ainet, tegi T. Eliot (1904).
Siiski kinnitasid keemilise sünapsi hüpoteesi lõpuks ainult G. Dale'i ja O. Löwy tööd. Dale tegi 1914. aastal kindlaks, et atsetüülkoliin jäljendab parasümpaatilise närvi ärritust. Löwy tõestas 1921. aastal, et atsetüülkoliin eraldub vagusnärvi närvilõpust ja 1926. aastal avastas ta atsetüülkoliinesteraasi, ensüümi, mis hävitab atsetüülkoliini.
Ergastus keemilises sünapsis edastatakse kasutades vahendaja. See protsess hõlmab mitut etappi. Vaatleme neid omadusi atsetüülkoliini sünapsi näitel, mis on laialt levinud kesknärvisüsteemis, autonoomses ja perifeerses närvisüsteemis (joonis 3).

Riis. 3. Keemilise sünapsi toimimise skeem
1. Vahendaja atsetüülkoliin (ACh) sünteesitakse sünaptilises naastudes atsetüül-CoA-st (atsetüülkoensüüm A moodustub mitokondrites) ja koliinist (sünteesitakse maksas), kasutades atsetüülkoliini transferaasi (joonis 3, 1).
2. Valik on sisse pakitud sünaptilised vesiikulid ( Castillo, Katz; 1955). Vahendaja kogus ühes vesiikulis on mitu tuhat molekuli ( vahendaja kvant). Mõned vesiikulid asuvad PreSM-il ja on valmis vahendaja vabastamiseks (joonis 3, 2).
3. Vahendaja vabastab eksotsütoos PreSM-i ergastamisel. Sissetulev vool mängib olulist rolli membraani purunemisel ja saatja kvantvabastamisel. Ca 2+ (Joon. 3, 3).
4. Vabastatud valik seondub spetsiifilise retseptorvalguga PostSM (joonis 3, 4).
5. Vahendaja ja retseptori interaktsiooni tulemusena ioonjuhtivuse muutused PostSM: kui Na + kanalid avanevad, depolarisatsioon; K + või Cl - kanalite avamine viib hüperpolarisatsioon(Joon. 3, 5).
6 . Pärast depolarisatsiooni käivituvad postsünaptilises tsütoplasmas biokeemilised protsessid (joonis 3, 6).
7. Retseptor vabaneb vahendajast: ACh hävitab atsetüülkoliinesteraas (AChE, joonis 3. 7).
Vormi algus
Pange tähele, et vahendaja suhtleb tavaliselt spetsiifilise retseptoriga teatud tugevuse ja kestusega. Miks on curare mürk? Kurare toimekoht on täpselt ACh sünaps. Curare seondub tugevamalt atsetüülkoliini retseptoriga ja jätab selle ilma interaktsioonist neurotransmitteriga (ACh). Ergastus somaatilistelt närvidelt skeletilihastesse, sealhulgas frenic närvist peamisse hingamislihasesse (diafragma) edastatakse ACh abil, seega põhjustab curare lihaste lõdvestamist ja hingamise seiskumist (mis tegelikult põhjustab surma).
Märgime peamist ergastuse ülekande tunnused keemilises sünapsis.
1. Ergastus edastatakse kasutades keemilist vahendajat – vahendajat.
2. Ergastus edastatakse ühes suunas: PreSm-st PostSm-i.
3. Keemilises sünapsis toimub ajutine viivitus ergastuse läbiviimisel, seetõttu on sünapsil madal labiilsus.
4. Keemiline sünaps on väga tundlik mitte ainult vahendajate, vaid ka teiste bioloogiliselt aktiivsete ainete, ravimite ja mürkide toime suhtes.
5. Keemilises sünapsis toimub ergastuste transformatsioon: ergastuse elektrokeemiline olemus PreSM-il jätkub sünaptiliste vesiikulite eksotsütoosi ja vahendaja seondumise spetsiifilise retseptoriga biokeemiliseks protsessiks. Sellele järgneb PostSM-i ioonjuhtivuse muutus (samuti elektrokeemiline protsess), mis jätkub biokeemiliste reaktsioonidega postsünaptilises tsütoplasmas.
Põhimõtteliselt peaks sellisel mitmeastmelisel ergastuse edastamisel olema märkimisväärne bioloogiline tähtsus. Pange tähele, et igal etapil on võimalik reguleerida ergastuse ülekande protsessi. Hoolimata vahendajate piiratud arvust (veidi rohkem kui tosin), on keemilises sünapsis tingimused sünapsi saabuva närviergastuse saatuse otsustamisel väga mitmekesised. Keemiliste sünapside tunnuste kombinatsioon selgitab närvi- ja vaimsete protsesside individuaalset biokeemilist mitmekesisust.
Nüüd peatume kahel olulisel postsünaptilises ruumis toimuval protsessil. Märkasime, et ACh interaktsiooni tulemusena PostSM-i retseptoriga võib areneda nii depolarisatsioon kui ka hüperpolarisatsioon. Mis määrab, kas vahendaja on ergastav või inhibeeriv? Vahendaja ja retseptori interaktsiooni tulemus määratakse retseptorvalgu omadustega(teine keemilise sünapsi oluline omadus on see, et PostSM on aktiivne sellele tuleva ergastuse suhtes). Põhimõtteliselt on keemiline sünaps dünaamiline moodustis, retseptorit muutes saab ergastuse saanud rakk mõjutada oma edasist saatust. Kui retseptori omadused on sellised, et selle interaktsioon saatjaga avab Na + kanalid, siis millal ühe vahendaja kvanti eraldamisega PostSM-is areneb lokaalne potentsiaal(neuromuskulaarse ristmiku jaoks nimetatakse seda miniatuurseks otsaplaadi potentsiaaliks - MEPP).
Millal PD tekib? PostSM-i ergastus (ergastav postsünaptiline potentsiaal – EPSP) tekib lokaalsete potentsiaalide liitmise tulemusena. Saate valida kahte tüüpi summeerimisprotsesse. Kell mitmete vahendajakvantide järjestikune vabastamine samas sünapsis(vesi kulutab kivi ära) tekib ajutineA Ma olen summeerimine. Kui kvantmediaatorid vabastatakse samaaegselt erinevates sünapsides(neuroni membraanil võib neid olla mitu tuhat) tekib ruumiline summeerimine. PostSM-i membraani repolariseerumine toimub aeglaselt ja pärast vahendaja üksikute kvantide vabanemist on PostSM mõnda aega kõrgendatud seisundis (nn sünaptiline potentsiatsioon, joonis 4). Võib-olla toimub sel viisil sünapsi treenimine (saatja kvantide vabanemine teatud sünapsides võib membraani "ette valmistada" otsustavaks interaktsiooniks saatjaga).
Kui PostSM-is avanevad K + või Cl - kanalid, ilmub inhibeeriv postsünaptiline potentsiaal (IPSP, joonis 4).

Riis. 4. Postsünaptilised membraanipotentsiaalid
Loomulikult saab IPSP arenemise korral ergastuse edasise levimise peatada. Teine võimalus ergastusprotsessi peatamiseks on presünaptiline inhibeerimine. Kui sünaptilise naastu membraanile moodustub inhibeeriv sünaps, võib PreSM-i hüperpolarisatsiooni tulemusena blokeerida sünaptiliste vesiikulite eksotsütoos.
Teine oluline protsess on biokeemiliste reaktsioonide areng postsünaptilises tsütoplasmas. PostSM-i ioonjuhtivuse muutus aktiveerib nn teisesed sõnumitoojad (vahendajad): cAMP, cGMP, Ca 2+ -sõltuv proteiinkinaas, mis omakorda aktiveerivad erinevaid proteiinkinaase neid fosforüülides. Need biokeemilised reaktsioonid võivad "laskuda" sügavale tsütoplasmasse kuni neuroni tuumani, reguleerides valgusünteesi protsesse. Seega saab närvirakk reageerida saabuvale ergastusele mitte ainult oma edasise saatuse üle otsustades (reageerida EPSP või IPSP-ga, st jätkata või mitte jätkata), vaid ka muuta retseptorite arvu või sünteesida retseptorvalku uuega. omadused seoses teatud vahendajaga. Järelikult veel üks oluline keemilise sünapsi omadus: tänu postsünaptilise tsütoplasma biokeemilistele protsessidele valmistub (õpib) rakk tulevasteks interaktsioonideks.
Närvisüsteemis toimivad mitmesugused sünapsid, mis erinevad vahendajate ja retseptorite poolest. Sünapsi nime määrab vahendaja, täpsemalt, konkreetse vahendaja retseptori nimetus. Seetõttu vaatleme peamiste närvisüsteemi vahendajate ja retseptorite klassifikatsiooni (vt ka loengus jagatavat materjali!!).
Oleme juba märkinud, et vahendaja ja retseptori vahelise interaktsiooni mõju määravad retseptori omadused. Seetõttu võivad tuntud vahendajad, välja arvatud g-aminovõihape, täita nii ergastavate kui ka inhibeerivate mediaatorite ülesandeid, mille keemilise struktuuri alusel eristatakse järgmisi vahendajate rühmi.
Atsetüülkoliin Kesknärvisüsteemis laialt levinud, on autonoomse närvisüsteemi kolinergiliste sünapside ja ka somaatiliste neuromuskulaarsete sünapside vahendaja (joonis 5).

Riis. 5. Atsetüülkoliini molekul
Teatud kahte tüüpi kolinergilised retseptorid: nikotiin ( H-kolinergilised retseptorid) ja muskariinid ( M-kolinergilised retseptorid). Nimetus anti ainetele, mis põhjustavad nendes sünapsides atsetüülkoliiniga sarnast toimet: N-kolinomimeetikum on nikotiin, A M-kolinomimeetikum- kärbseseene toksiin Amanita muscaria ( muskariin). H-kolinergilise retseptori blokaator (antikolinergiline) on d-tubokurariin(kuraremürgi põhikomponent) ja M-antikolinergiline on Atropa belladonna belladonna toksiin – atropiin. Huvitav on see, et atropiini omadused on juba ammu teada ja oli aeg, mil naised kasutasid belladonnast saadud atropiini nägemispupillide laienemiseks (silmade tumedaks ja “ilusamaks muutmiseks”).
Järgmisel neljal peamisel vahendajal on keemilise struktuuri sarnasused, seega klassifitseeritakse need järgmiselt monoamiinid. See serotoniin või 5-hüdroksütrüptamiinid (5-HT), mängivad olulist rolli tugevdamise mehhanismides (rõõmuhormoon). Seda sünteesitakse inimese jaoks asendamatust aminohappest – trüptofaanist (joonis 6).

Riis. 6. Serotoniini (5-hüdroksütrüptamiini) molekul
Kolm muud vahendajat sünteesitakse asendamatust aminohappest fenüülalaniinist ja on seetõttu ühendatud üldnimetuse all. katehhoolamiinid- See dopamiin (dopamiin), norepinefriin (norepinefriin) ja adrenaliin (epinefriin, joonis 7).

Riis. 7. Katehhoolamiinid
hulgas aminohapped vahendajate hulka gamma-aminovõihape(g-AMK või GABA – tuntud kui ainus inhibeeriv neurotransmitter), glütsiin, glutamiinhape, asparagiinhape.
Vahendajad hõlmavad mitmeid peptiidid. 1931. aastal avastas Euler aju- ja soolteekstraktidest aine, mis põhjustab soolestiku silelihaste kokkutõmbumist ja veresoonte laienemist. See saatja eraldati puhtal kujul hüpotalamusest ja sai nime aine P(inglise keelest pulber - pulber, koosneb 11 aminohappest). Hiljem tehti kindlaks, et ainel P on oluline roll valulike ergastuste läbiviimisel (nimetust ei pidanud muutma, kuna valu inglise keeles on valu).
Delta une peptiid sai oma nime võime tõttu tekitada elektroentsefalogrammis aeglaseid kõrge amplituudiga rütme (deltarütme).
Ajus sünteesitakse mitmeid narkootilisi (opiaate) valgu vahendajaid. Need on pentapeptiidid Met-enkefaliin Ja Leu-enkefaliin ja endorfiine. Need on olulisemad valu ergutuste blokaatorid ja tugevdamise (rõõmu ja naudingu) vahendajad. Teisisõnu, meie aju on suurepärane tehas endogeenne ravimid. Peamine on õpetada aju neid tootma. "Kuidas?" - te küsite. See on lihtne – endogeensed opiaadid tekivad siis, kui kogeme naudingut. Tehke kõike mõnuga, sundige oma endogeenset tehast opiaate sünteesima! See võimalus on meile loomulikult antud sünnist saati – valdav enamus neuroneid reageerivad positiivsele tugevdusele.
Viimaste aastakümnete uuringud on võimaldanud avastada veel ühe väga huvitava vahendaja - lämmastikoksiid (NO). Selgus, et NO ei mängi mitte ainult olulist rolli veresoonte toonuse reguleerimisel (teile teadaolev nitroglütseriin on NO allikas ja laiendab koronaarsooni), vaid sünteesitakse ka kesknärvisüsteemi neuronites.
Põhimõtteliselt pole vahendajate ajalugu veel lõppenud, närvilise ergastuse reguleerimises osalevad mitmed ained. Lihtsalt nende sünteesi fakt neuronites ei ole veel täpselt kindlaks tehtud, neid pole leitud sünaptilistest vesiikulitest ega leitud neile spetsiifilisi retseptoreid.
Föderaalne Haridusagentuur
Riiklik õppeasutus
erialane kõrgharidus
"Ryazani osariigi ülikool, mis sai nime S.A. Yesenin"
Psühholoogia, Pedagoogika ja Sotsiaaltöö Instituut
Testitöö erialal "Neurofüsioloogia ja VND põhialused"
teemal: „Sünapsi mõiste, sünapsi struktuur.
Ergastuse ülekanne sünapsis"
Lõpetanud 13L rühma õpilane
1. aasta OZO (3) A.I. Šarova
Kontrollitud:
meditsiiniteaduste professor
O.A. Belova
Rjazan 2010
1. Sissejuhatus………………………………………………………………..3
2. Sünapsi struktuur ja funktsioonid……………………………………………6
3. Ergastuse ülekanne sünapsis……………………………………….8
4. Keemiline sünaps………………………………………………………………9
5. Vahendaja isoleerimine…………………………………………………10
6. Keemilised vahendajad ja nende liigid………………………………..12
7. Järeldus………………………………………………………………15
8. Viidete loetelu……………………………………………………………………..17
Sissejuhatus.
Meie keha on üks suur kellamehhanism. See koosneb suurest hulgast pisikestest osakestest, mis asuvad selles ranges järjekorras ja igaüks neist täidab teatud funktsioone ja omab oma ainulaadsed omadused. See mehhanism - keha - koosneb rakkudest, mis ühendavad nende kudesid ja süsteeme: kõik see tervikuna kujutab endast üht ahelat, keha supersüsteemi. Suurim hulk rakulisi elemente ei saaks töötada ühtse tervikuna, kui kehas ei eksisteeriks keerulist regulatsioonimehhanismi. Närvisüsteem mängib regulatsioonis erilist rolli. Kogu närvisüsteemi kompleksne töö – siseorganite töö reguleerimine, liigutuste juhtimine, olgu siis lihtsad ja teadvustamata liigutused (näiteks hingamine) või inimese keerukad käteliigutused – kõik see põhineb sisuliselt rakud üksteisega. Kõik see põhineb sisuliselt signaali edastamisel ühest rakust teise. Lisaks täidab iga rakk oma tööd ja mõnikord on tal mitu funktsiooni. Funktsioonide mitmekesisust pakuvad kaks tegurit: viis, kuidas rakud on üksteisega ühendatud, ja viis, kuidas need ühendused on paigutatud. Ergastuse üleminek (ülekanne) närvikiust rakku, mida see innerveerib (närv, lihas, sekretoorne) toimub spetsiaalse moodustumise kaudu, mida nimetatakse sünapsiks.
Sünapsi struktuur ja funktsioonid.
Iga hulkrakne organism, iga rakkudest koosnev kude vajab mehhanisme, mis tagavad rakkudevahelise interaktsiooni. Vaatame, kuidas neid teostatakse interneuronaalneinteraktsioonid. Info liigub vormis mööda närvirakku tegevuspotentsiaalid. Ergastuse ülekandmine aksoni otstest innerveeritud elundisse või muusse närvirakku toimub rakkudevaheliste struktuursete moodustiste kaudu - sünapsid (kreeka keelest "Synapsis" - ühendus, ühendus). Sünapsi mõiste võttis kasutusele inglise füsioloog C. Sherrington aastal 1897, tähistamaks neuronite vahelist funktsionaalset kontakti. Tuleb märkida, et juba eelmise sajandi 60ndatel NEED. Sechenov rõhutas, et ilma rakkudevahelise suhtluseta on võimatu selgitada isegi kõige elementaarsema närviprotsessi tekkemeetodeid. Mida keerulisem on närvisüsteem ja mida suurem on neuraalsete ajuelementide arv, seda olulisemaks muutub sünaptiliste kontaktide tähtsus.
Erinevad sünaptilised kontaktid erinevad üksteisest. Kuid sünapside mitmekesisuse juures on nende struktuuril ja funktsioonil teatud ühised omadused. Seetõttu kirjeldame esmalt nende toimimise üldpõhimõtteid.
Sünaps - on keeruline struktuurne moodustis, mis koosneb
presünaptiline membraan - elektrogeenne membraan aksoni terminalis, moodustab lihasraku sünapsi (enamasti on see aksoni terminaalne haru)
postsünaptiline membraan - innerveeritud raku elektrogeenne membraan, millel moodustub sünaps (enamasti on see kehamembraani osa või teise neuroni dendriit)
sünaptiline lõhe - presünaptilise ja postsünaptilise membraani vaheline ruum, täidetud vedelikuga, mis koostiselt meenutab vereplasma
Sünapsid võivad asuda kahe neuroni vahel (interneuronaalne), neuronite ja lihaskiudude vahel (neuromuskulaarne) retseptori moodustiste ja sensoorsete neuronite protsesside vahel (retseptor-neuronaalne), neuroniprotsesside ja teiste rakkude vahel ( näärmeline).
Sünapside klassifikatsioone on mitu.
1. Lokaliseerimise järgi:
1) tsentraalsed sünapsid;
2) perifeersed sünapsid.
Kesksünapsid asuvad kesknärvisüsteemis ja neid leidub ka autonoomse närvisüsteemi ganglionides.
Tsentraalsed sünapsid– need on kontaktid kahe närviraku vahel ja need kontaktid on heterogeensed ja olenevalt struktuurist, millel esimene neuron moodustab teise neuroniga sünapsi, eristatakse neid:
a) aksosomaatiline, moodustatud ühe neuroni aksonist ja teise neuroni kehast;
b) aksodendriit, mille moodustavad ühe neuroni akson ja teise neuroni dendriit;
c) aksoaksonaalne (esimese neuroni akson moodustab teise neuroni aksonil sünapsi);
d) dendrodentriit (esimese neuroni dendriit moodustab teise neuroni dendriidil sünapsi).
Neid on mitut tüüpi perifeersed sünapsid:
a) müoneuraalne (neuromuskulaarne), mille moodustavad motoorse neuroni ja lihasraku akson;
b) neuroepiteliaalne, moodustatud neuroni ja sekretoorse raku aksonist.
2. Sünapside funktsionaalne klassifikatsioon:
1) ergastavad sünapsid;
2) inhibeerivad sünapsid.
ergastav sünaps- sünaps, milles postsünaptiline membraan on ergastatud; selles tekib ergastav postsünaptiline potentsiaal ja sünapsi saabuv erutus levib edasi.
Inhibeeriv sünaps- A. Sünaps, mille postsünaptilisel membraanil tekib inhibeeriv postsünaptiline potentsiaal ja sünapsi saabuv erutus ei levi edasi; B. ergastav aksoaksonaalne sünaps, mis põhjustab presünaptilist inhibeerimist.
3. Vastavalt sünapsi ergastuse ülekande mehhanismidele:
1) keemiline;
2) elektriline;
3) segatud
Omapära keemilised sünapsid seisneb selles, et ergastuse ülekandmine toimub spetsiaalse kemikaalide rühma abil - vahendajad. See on rohkem spetsialiseerunud kui elektriline sünaps.
Neid on mitut tüüpi keemilised sünapsid olenevalt vahendaja iseloomust:
a) kolinergiline.
b) adrenergilised.
c) dopamiinergiline. Nad edastavad põnevust dopamiini abil;
d) histaminergiline. Nad edastavad erutust histamiini abil;
e) GABAergiline. Nendes edastatakse erutus gamma-aminovõihappe abil, st areneb inhibeerimisprotsess.
Adrenergiline sünaps - sünaps, mille vahendajaks on norepinefriin. See edastab ergastuse kolme katehhoolamiini abil; Seal on a1-, b1- ja b2-adrenergilised sünapsid. Need moodustavad sümpaatilise närvisüsteemi neuroorganite sünapsid ja kesknärvisüsteemi sünapsid. a-adrenoreaktiivsete sünapside ergastamine põhjustab vasokonstriktsiooni ja emaka kokkutõmbumist; b1- adrenoreaktiivsed sünapsid - suurenenud südamefunktsioon; b2 - adrenoreaktiivne - bronhide laienemine.
Kolinergiline sünaps - selles sisalduv vahendaja on atsetüülkoliin. Need jagunevad n-kolinergilisteks ja m-kolinergilisteks sünapsideks.
M-kolinergilises Sünapsis on postsünaptiline membraan muskariini suhtes tundlik. Need sünapsid moodustavad parasümpaatilise süsteemi neuroorganite sünapsid ja kesknärvisüsteemi sünapsid.
N-kolinergilises Sünapsis on postsünaptiline membraan tundlik nikotiini suhtes. Seda tüüpi sünapsid moodustavad somaatilise närvisüsteemi neuromuskulaarsed sünapsid, ganglionide sünapsid, sümpaatilise ja parasümpaatilise närvisüsteemi sünapsid ning kesknärvisüsteemi sünapsid.
Elektriline sünaps- selles kandub erutus pre- postsünaptilisele membraanile elektriliselt, s.o. toimub ergastuse efaptiline ülekanne – aktsioonipotentsiaal jõuab presünaptilisse terminali ja levib seejärel rakkudevaheliste kanalite kaudu, põhjustades postsünaptilise membraani depolarisatsiooni. Elektrilises sünapsis saatjat ei toodeta, sünaptiline lõhe on väike (2 - 4 nm) ja seal on 1 - 2 nm laiused valgu sillad-kanalid, mida mööda liiguvad ioonid ja väikesed molekulid. See aitab kaasa madalale postsünaptilisele membraanikindlusele. Seda tüüpi sünapsid on palju vähem levinud kui keemilised sünapsid ja erinevad neist suurema ergastuse ülekande kiiruse, kõrge töökindluse ja ergastuse kahesuunalise juhtivuse võimaluse poolest.
Sünapsitel on mitmeid füsioloogilisi omadusi :
1) sünapside klapiomadus, st võime edastada ergastust ainult ühes suunas presünaptilisest membraanist postsünaptilisse;
2) sünaptilise viivituse omadus, mis on tingitud asjaolust, et ergastuse ülekande kiirus väheneb;
3) võimendusomadus(iga järgnev impulss viiakse läbi lühema postsünaptilise viivitusega). See on tingitud asjaolust, et eelmise impulsi saatja jääb presünaptilisele ja postsünaptilisele membraanile;
4) madal sünapsi labiilsus(100–150 impulssi sekundis).
Ergastuse ülekanne sünapsis.
Sünapside ülekandumise mehhanism jäi pikka aega ebaselgeks, kuigi oli ilmne, et signaali edastamine sünaptilises piirkonnas erineb järsult aktsioonipotentsiaali läbiviimisest piki aksonit. 20. sajandi alguses püstitati aga hüpotees, et sünaptiline ülekanne toimub kas elektriline või keemiliselt. Sünaptilise ülekande elektriteooriat kesknärvisüsteemis tunnustati kuni 50. aastate alguseni, kuid see kaotas märkimisväärselt pärast seda, kui mitmel juhul demonstreeriti keemilist sünapsi. perifeersed sünapsid. Näiteks, A.V. Kibjakov, Pärast närviganglioni eksperimendi läbiviimist, samuti mikroelektrooditehnoloogia kasutamist kesknärvisüsteemi neuronite sünaptilise potentsiaali rakusiseseks registreerimiseks, oli võimalik teha järeldus ülekande keemilise olemuse kohta seljaaju interneuronaalsetes sünapsides.
Viimaste aastate mikroelektroodiuuringud on näidanud, et teatud interneuronite sünapsides eksisteerib elektriline ülekandemehhanism. Nüüd on ilmnenud, et on olemas nii keemilise kui ka elektrilise ülekandemehhanismiga sünapsid. Pealegi toimivad mõnes sünaptilises struktuuris koos nii elektrilised kui keemilised ülekandemehhanismid – need on nn segatud sünapsid.
Kui primitiivsemate loomade närvisüsteemile on iseloomulikud elektrilised sünapsid (koelenteraatide närvidifusioonisüsteem, jõevähkide ja anneliidide mõned sünapsid, kalade närvisüsteemi sünapsid), kuigi neid leidub imetajate ajus. Kõigil ülalnimetatud juhtudel edastatakse impulsse läbi depolariseeriv presünaptilises elemendis tekkiva elektrivoolu toime. Samuti tahaksin märkida, et elektriliste sünapside puhul on impulsi edastamine võimalik nii ühes kui ka kahes suunas. Ka madalamatel loomadel kontakt vahel presünaptiline Ja postsünaptiline element viiakse läbi ainult ühe sünapsi kaudu - monosünaptiline suhtlusvorm, fülogeneesi protsessis toimub aga üleminek sellele polüsünaptiline suhtlusvorm, see tähendab, kui ülaltoodud kontakt toimub suurema hulga sünapside kaudu.
Käesolevas töös tahaksin aga pikemalt peatuda keemilise ülekandemehhanismiga sünapsitel, mis moodustavad suurema osa kõrgemate loomade ja inimeste kesknärvisüsteemi sünaptilisest aparaadist. Seega on keemilised sünapsid minu arvates eriti huvitavad, kuna need pakuvad väga keerulisi rakkude interaktsioone ja on seotud ka mitmete patoloogiline protsessid ja muuta nende omadusi teatud ravimite mõju all.
Lihas- ja näärmerakud edastatakse spetsiaalse struktuurse moodustumise - sünapsi kaudu.
Sünaps- struktuur, mis tagab signaali juhtimise ühelt teisele. Selle termini võttis kasutusele inglise füsioloog C. Sherrington 1897. aastal.
Sünapsi struktuur
Sünapsid koosnevad kolmest põhielemendist: presünaptilisest membraanist, postsünaptilisest membraanist ja sünaptilisest lõhest (joonis 1).
Riis. 1. Sünapsi struktuur: 1 - mikrotuubulid; 2 - mitokondrid; 3 - saatjaga sünaptilised vesiikulid; 4 - presünaptiline membraan; 5 - postsünaptiline membraan; 6 - retseptorid; 7 - sünaptiline lõhe
Mõnel sünapsi elemendil võivad olla teised nimed. Näiteks sünaptiline naast on sünaps nende vahel, otsaplaat on postsünaptiline membraan, motoorne tahvel on aksoni presünaptiline ots lihaskiul.

Presünaptiline membraan katab laienenud närvilõpme, mis on neurosekretoorne aparaat. Presünaptiline osa sisaldab vesiikuleid ja mitokondreid, mis tagavad vahendaja sünteesi. Vahendajad ladestuvad graanulitena (mullides).
Postsünaptiline membraan - rakumembraani paksenenud osa, millega presünaptiline membraan on kontaktis. Sellel on ioonkanalid ja see on võimeline tekitama aktsioonipotentsiaali. Lisaks sisaldab see spetsiaalseid valgu struktuure - retseptoreid, mis tajuvad vahendajate toimet.
Sünaptiline lõhe on ruum presünaptiliste ja postsünaptiliste membraanide vahel, mis on täidetud sarnase koostisega vedelikuga.

Riis. Sünapsi struktuur ja sünaptilise signaali edastamise käigus läbiviidavad protsessid
Sünapside tüübid
Sünapsid liigitatakse asukoha, toime laadi ja signaali edastamise meetodi järgi.
Asukoha järgi Nad eristavad neuromuskulaarseid sünapse, neuroglandulaarseid ja neuroneuronaalseid; viimased omakorda jagunevad aksoaksonaalseteks, aksodendriitilisteks, aksosomaatiliseks, dendrosomaatiliseks, dendrodendrootilisteks.
Tegevuse olemuse järgi Pertseptiivse struktuuri sünapsid võivad olla ergutavad või inhibeerivad.
Signaali edastamise meetodil Sünapsid jagunevad elektrilisteks, keemilisteks ja segatud.
Tabel 1. Sünapside klassifikatsioon ja liigid

Sünapside klassifikatsioon ja ergastuse ülekandemehhanism
Sünapsid liigitatakse järgmiselt:
- asukoha järgi - perifeerne ja keskne;
- oma tegevuse olemuse järgi - põnev ja pärssiv;
- signaali edastamise meetodil - keemiline, elektriline, segatud;
- vastavalt vahendajale, mille kaudu ülekanne toimub - kolinergiline, adrenergiline, serotonergiline jne.
Põnevus kandub edasi vahendajad(vahendajad).
Vahendajad- keemiliste ainete molekulid, mis tagavad ergastuse edasikandumise sünapsides. Teisisõnu, keemilised ained, mis osalevad ergastuse või inhibeerimise ülekandmisel ühest erutuvast rakust teise.
Vahendajate omadused
- Sünteesitakse neuronis
- Kogunevad raku lõppu
- Vabaneb, kui Ca2+ ioon ilmub presünaptilisse terminali
- Omavad spetsiifilist mõju postsünaptilisele membraanile
Vastavalt keemilisele struktuurile võib vahendajad jagada amiinideks (norepinefriin, dopamiin, serotoniin), aminohapeteks (glütsiin, gamma-aminovõihape) ja polüpeptiidideks (endorfiinid, enkefaliinid). Atsetüülkoliini tuntakse peamiselt ergastava neurotransmitterina ja seda leidub kesknärvisüsteemi erinevates osades. Saatja asub presünaptilise paksenemise (sünaptilise naastu) vesiikulites. Vahendaja sünteesitakse neuronirakkudes ja seda saab uuesti sünteesida sünaptilises lõhes lõhustunud metaboliitidest.
Kui aksoni terminalid on ergastatud, sünaptilise naastu membraan depolariseerub, põhjustades kaltsiumiioonide voolamist rakuvälisest keskkonnast närvilõpmesse kaltsiumikanalite kaudu. Kaltsiumiioonid stimuleerivad sünaptiliste vesiikulite liikumist presünaptilisele membraanile, nende sulandumist sellega ja sellele järgnevat saatja vabanemist sünaptilisse pilusse. Pärast pilusse tungimist difundeerub saatja postsünaptilisse membraani, mis sisaldab selle pinnal retseptoreid. Saatja interaktsioon retseptoritega põhjustab naatriumikanalite avanemist, mis aitab kaasa postsünaptilise membraani depolarisatsioonile ja ergastava postsünaptilise potentsiaali ilmnemisele. Neuromuskulaarses sünapsis nimetatakse seda potentsiaali otsaplaadi potentsiaal. Depolariseeritud postsünaptilise membraani ja sama membraani külgnevate polariseeritud osade vahel tekivad lokaalsed voolud, mis depolariseerivad membraani kriitilise tasemeni, millele järgneb aktsioonipotentsiaali teke. Aktsioonipotentsiaal levib näiteks lihaskiu kõigis membraanides ja põhjustab selle kokkutõmbumise.
Sünaptilisse pilusse vabanev saatja seondub postsünaptilise membraani retseptoritega ja lõhustub vastava ensüümi toimel. Seega hävitab koliinesteraas neurotransmitteri atsetüülkoliini. Pärast seda siseneb teatud kogus vahendajate lagunemissaadusi sünaptilisse naastu, kus neist taassünteesitakse atsetüülkoliin.
Keha ei sisalda mitte ainult ergastavaid, vaid ka inhibeerivaid sünapse. Ergastuse ülekandemehhanismi järgi on need sarnased ergastavate sünapsidega. Inhibeerivates sünapsides seostub saatja (näiteks gamma-aminovõihape) postsünaptilise membraani retseptoritega ja soodustab selle avanemist. Sel juhul aktiveeritakse nende ioonide tungimine rakku ja areneb postsünaptilise membraani hüperpolarisatsioon, mis põhjustab inhibeeriva postsünaptilise potentsiaali ilmnemist.
Nüüdseks on leitud, et üks vahendaja võib seonduda mitme erineva retseptoriga ja kutsuda esile erinevaid reaktsioone.
Keemilised sünapsid
Keemiliste sünapside füsioloogilised omadused
Ergastuse keemilise ülekandega sünapsitel on teatud omadused:
- ergastamine toimub ühes suunas, kuna saatja vabaneb ainult sünaptilisest naastust ja interakteerub postsünaptilise membraani retseptoritega;
- ergastuse levik sünapside kaudu toimub aeglasemalt kui piki närvikiudu (sünaptiline viivitus);
- ergastuse edastamine toimub spetsiaalsete vahendajate abil;
- ergastuse muutuste rütm sünapsides;
- sünapsid võivad väsida;
- sünapsid on väga tundlikud erinevate kemikaalide ja hüpoksia suhtes.
Ühesuunaline signaaliedastus. Signaal edastatakse ainult presünaptiliselt membraanilt postsünaptilisse membraani. See tuleneb sünaptiliste struktuuride struktuurilistest iseärasustest ja omadustest.
Aeglane signaaliedastus. Põhjuseks sünaptiline viivitus signaali edastamisel ühest rakust teise. Viivituse põhjuseks on saatja vabanemisprotsesside jaoks kuluv aeg, selle difusioon postsünaptilisele membraanile, postsünaptilise membraani retseptoritega seondumine, depolarisatsioon ja postsünaptilise potentsiaali muundamine AP-ks (aktsioonipotentsiaal). Sünaptilise viivituse kestus on vahemikus 0,5 kuni 2 ms.
Võimalus võtta kokku sünapsi saabuvate signaalide mõju. See summeerimine ilmub siis, kui järgnev signaal saabub sünapsi veidi aega (1-10 ms) pärast eelmist. Sellistel juhtudel suureneb EPSP amplituud ja postsünaptilisel neuronil võib tekkida suurem AP sagedus.
Põnevuse rütmi ümberkujundamine. Presünaptilisele membraanile saabuvate närviimpulsside sagedus ei vasta tavaliselt postsünaptilise neuroni poolt genereeritud AP-de sagedusele. Erandiks on sünapsid, mis edastavad ergastuse närvikiust skeletilihastele.
Sünapside madal labiilsus ja suur väsimus. Sünapsid võivad läbi viia 50-100 närviimpulsi sekundis. See on 5-10 korda väiksem kui maksimaalne AP sagedus, mida närvikiud suudavad elektrilisel stimuleerimisel reprodutseerida. Kui närvikiude peetakse praktiliselt väsimatuks, siis sünapsi ajal tekib väsimus väga kiiresti. See ilmneb saatja reservide, energiaressursside ammendumise, postsünaptilise membraani püsiva depolarisatsiooni jne tõttu.
Sünapside kõrge tundlikkus bioloogiliselt aktiivsete ainete, ravimite ja mürkide toime suhtes. Näiteks mürk strühniin blokeerib kesknärvisüsteemi inhibeerivate sünapside funktsiooni, seondudes vahendaja glütsiini suhtes tundlike retseptoritega. Teetanuse toksiin blokeerib inhibeerivad sünapsid, häirides saatja vabanemist presünaptilisest terminalist. Mõlemal juhul arenevad välja eluohtlikud nähtused. Näiteid bioloogiliselt aktiivsete ainete ja mürkide mõjust signaaliülekandele neuromuskulaarsetes sünapsides on käsitletud eespool.
Sünoptilise ülekande hõlbustamise ja depressiooni omadused. Sünaptilise ülekande hõlbustamine toimub siis, kui närviimpulsid jõuavad sünapsi lühikese aja (10-50 ms) järel üksteise järel, s.o. piisavalt sageli. Pealegi põhjustab iga järgnev presünaptilisele membraanile saabuv PD teatud aja jooksul saatja sisalduse suurenemist sünaptilises pilus, EPSP-de amplituudi suurenemist ja sünaptilise ülekande efektiivsuse tõusu.
Üks soodustamise mehhanisme on Ca 2 ioonide akumuleerumine presünaptilisse terminali. Kaltsiumipumbal kulub mitukümmend millisekundit, et eemaldada kaltsiumi osa, mis sisenes AP saabumisel sünaptilisse terminali. Kui sel ajal saabub uus aktsioonipotentsiaal, siseneb terminali uus osa kaltsiumi ja selle mõju neurotransmitteri vabanemisele lisatakse kaltsiumi jääkkogusele, mida kaltsiumipumbal ei olnud aega neuroplasmast eemaldada. terminali.
Leevenduse arendamiseks on ka teisi mehhanisme. Seda nähtust nimetatakse ka klassikalistes füsioloogiaõpikutes post-teetaniline võimendamine. Sünaptilise ülekande hõlbustamine on oluline mälumehhanismide toimimises, konditsioneeritud reflekside kujunemisel ja õppimisel. Signaali edastamise hõlbustamine on sünaptilise plastilisuse arendamise ja nende funktsioonide täiustamise aluseks sagedase aktiveerimisega.
Sünapsi signaaliülekande depressioon (inhibeerimine) tekib siis, kui presünaptilisele membraanile saabuvad väga sagedased (neuromuskulaarse sünapsi puhul üle 100 Hz) närviimpulsid. Depressiooni nähtuse arengumehhanismides on saatja reservide ammendumine presünaptilises terminalis, postsünaptilise membraani retseptorite tundlikkuse vähenemine saatja suhtes ja postsünaptilise membraani püsiva depolarisatsiooni areng, mis raskendab genereerimist. postsünaptilise raku membraanil olevad AP-d on olulised.
Elektrilised sünapsid
Lisaks ergastuse keemilise ülekandega sünapsidele on kehal elektriülekandega sünapsid. Nendel sünapsitel on väga kitsas sünaptiline lõhe ja kahe membraani vahel vähenenud elektritakistus. Membraanidevaheliste põikkanalite olemasolu ja madala takistuse tõttu läbib membraane kergesti elektriline impulss. Elektrilised sünapsid on tavaliselt iseloomulikud sama tüüpi rakkudele.
Stiimuliga kokkupuute tulemusena ergastab presünaptiline aktsioonipotentsiaal postsünaptilist membraani, kus tekib leviv aktsioonipotentsiaal.
Neid iseloomustab suurem ergastuskiirus võrreldes keemiliste sünapsidega ja madal tundlikkus kemikaalide mõjude suhtes.
Elektrilistel sünapsidel on ergastuse ühe- ja kahesuunaline ülekanne.
Kehas leidub ka elektrilisi inhibeerivaid sünapse. Inhibeeriv toime areneb välja voolu toimel, mis põhjustab postsünaptilise membraani hüperpolarisatsiooni.
Segasünapsides saab ergastuse edastada nii elektriliste impulsside kui ka vahendajate abil.
- Kokkupuutel 0
- Google+ 0
- Okei 0
- Facebook 0






