Ulančano nasljeđe - nasljeđivanje osobina čiji su geni lokalizirani na jednom kromosomu. Snaga povezanosti između gena ovisi o njihovoj udaljenosti: što su geni udaljeni jedan od drugoga, veća je učestalost križanja i obrnuto. Uz svojstva koja se nasljeđuju samostalno, moraju postojati i ona koja se nasljeđuju vezano jer su određena genima koji se nalaze na istom kromosomu. Takvi geni nastaju grupa kvačila. Broj veznih skupina u organizmima određene vrste jednak je broju kromosoma u haploidnom setu (npr. kod Drosophile 1 par = 4, kod čovjeka 1 par = 23).
Potpuni stisak- tip vezanog nasljeđivanja u kojem su geni analiziranih svojstava smješteni tako blizu jedni drugima da križanje između njih postaje nemoguće.
Nepotpuno kvačilo- tip vezanog nasljeđivanja u kojem se geni analiziranih svojstava nalaze na određenoj udaljenosti jedan od drugog, što omogućuje njihovo križanje.
(Crossover gamete- gamete u procesu formiranja kojih je došlo do crossingovera. Crossover gamete u pravilu čine mali dio ukupnog broja gameta.
Prelazak preko- izmjena dijelova homolognih kromosoma tijekom stanične diobe, uglavnom u profazi prve mejotske diobe, ponekad u mitozi. Pokusi T. Morgana, K. Bridgesa i A. Sturtevanta pokazali su da ne postoji apsolutno potpuna povezanost gena, u kojoj bi se geni uvijek zajedno prenosili. Vjerojatnost da se dva gena smještena na istom kromosomu neće razdvojiti tijekom mejoze kreće se od 1 do 0,5. U prirodi prevladava nepotpuno povezivanje, uzrokovano presjekom homolognih kromosoma i rekombinacijom gena. Citološku sliku crossing overa prvi je opisao danski znanstvenik F. Janssens.
Crossing over se događa samo kada su geni u heterozigotnom stanju (AB/av). Ako su geni u homozigotnom stanju (AB/AB ili aB/aB), izmjena identičnih dijelova ne daje nove kombinacije gena u gametama iu generaciji. Učestalost (postotak) crossing overa između gena ovisi o njihovoj udaljenosti: što su udaljeniji jedan od drugoga, to se crossing over češće događa. T. Morgan predložio je mjerenje udaljenosti između gena crossing overom kao postotak, koristeći formulu:
N1/N2 X 100 = % križanja,
gdje je N1 ukupan broj jedinki u F;
N2 je ukupan broj križanih jedinki.
Segment kromosoma na kojem se dogodi 1% crossing overa jednak je jednom morganidu (konvencionalna mjera udaljenosti između gena). Frekvencija križanja koristi se za određivanje relativnog položaja gena i udaljenosti između njih. Nove tehnologije koriste se za izradu ljudske genetske karte, a također su izrađene i citogenetske karte kromosoma.
Postoji nekoliko vrsta križanja: dvostruko, višestruko (složeno), nepravilno, neravnomjerno.
Crossing over dovodi do nove kombinacije gena i uzrokuje promjenu fenotipa. Osim toga, uz mutacije, važan je čimbenik u evoluciji organizama.)
Rezultat istraživanja T. Morgana bio je njegovo stvaranje kromosomske teorije nasljeđa:
· geni se nalaze na kromosomima; različiti kromosomi sadrže različit broj gena; skup gena svakog od nehomolognih kromosoma je jedinstven;
· svaki gen ima određeno mjesto (lokus) na kromosomu; alelni geni nalaze se u identičnim lokusima homolognih kromosoma;
· geni su smješteni na kromosomima u određenom linearnom nizu;
· geni lokalizirani na istom kromosomu nasljeđuju se zajedno, tvoreći vezu; broj veznih skupina jednak je haploidnom skupu kromosoma i konstantan je za svaku vrstu organizma;
· povezivanje gena može se poremetiti tijekom crossing overa, što dovodi do stvaranja rekombinantnih kromosoma; učestalost crossing overa ovisi o udaljenosti između gena: što je veća udaljenost, veća je veličina crossing overa;
· Svaka vrsta ima jedinstveni set kromosoma - kariotip.
Nasljeđivanje spola i spolno vezanih svojstava. Spolni kromosomi i njihova uloga u određivanju spola. Nasljeđe spola. Spol jedinke složena je osobina, nastala kako djelovanjem gena tako i razvojnim uvjetima. Ljudi imaju jedan od 23 para kromosoma - spolne kromosome, označene X i Y. Žene su homogametni spol, t.j. imaju dva X kromosoma, jedan od majke, a drugi od oca. Muškarci su heterogametni spol, imaju jedan X i jedan Y kromosom, pri čemu se X prenosi od majke, a Y od oca. Imajte na umu da heterogametni spol nije uvijek nužno muški; npr. kod ptica su ženke, dok su mužjaci homogametni. Postoje i drugi mehanizmi određivanja spola. Dakle, u određenom broju insekata Y kromosom je odsutan. U ovom slučaju, jedan od spolova razvija se u prisutnosti dva X kromosoma, a drugi - u prisutnosti jednog X kromosoma. Kod nekih kukaca spol je određen omjerom broja autosoma i spolnih kromosoma. Kod niza životinja tzv ponovno određivanje spola, kada se, ovisno o okolišnim čimbenicima, zigota razvija ili u žensko ili u muško. Razvoj spola kod biljaka ima iste raznolike genetske mehanizme kao i kod životinja.
Osobine povezane s X kromosomom. Ako se gen nalazi na spolnom kromosomu (naziva se spolno vezan), tada njegova manifestacija u potomcima slijedi drugačija pravila nego kod autosomnih gena. Pogledajmo gene koji se nalaze na X kromosomu. Kći nasljeđuje dva X kromosoma: jedan od majke i jedan od oca. Sin ima samo jedan X kromosom – od majke; Od oca dobiva Y kromosom. Dakle, otac gene na svom kromosomu X prenosi samo kćeri, ali ih sin ne može primiti. Budući da je kromosom X "bogatiji" genima u usporedbi s kromosomom Y, u tom smislu kći je genetski sličnija ocu nego sinu; sin je sličniji majci nego ocu.
Jedna od povijesno najpoznatijih karakteristika povezanih sa spolom kod ljudi je hemofilija, koja dovodi do teškog krvarenja od najmanjih posjekotina i opsežnih hematoma od modrica. Uzrokuje ga recesivni defektni alel 0, koji blokira sintezu proteina potrebnog za zgrušavanje krvi. Gen za ovaj protein je lokaliziran na X kromosomu. Heterozigotna žena +0 (+ znači normalan aktivni alel, dominantan u odnosu na alel 0 hemofilije) ne razvija hemofiliju, kao ni njezine kćeri, osim ako otac ima ovu patologiju. Međutim, njezin sin može dobiti alel 0 i zatim razviti hemofiliju. Recesivne bolesti uzrokovane genima na kromosomu X znatno rjeđe pogađaju žene nego muškarce, budući da se kod njih bolest očituje samo u slučajevima homozigotnosti - prisutnosti recesivnog alela u svakom od dva homologna kromosoma X; muškarci su pogođeni kad god njihov jedini X kromosom nosi defektni alel.
Veza s Y kromosomom.Informacije o genima koji se nalaze na Y kromosomu vrlo su oskudne. Pretpostavlja se da praktički ne nosi gene koji određuju sintezu proteina potrebnih za funkcioniranje stanice. Ali igra ključnu ulogu u razvoju muškog fenotipa. Odsutnost Y kromosoma u prisutnosti samo jednog X kromosoma dovodi do tzv. Turnerov sindrom: razvoj ženskog fenotipa sa slabo razvijenim primarnim i sekundarnim spolnim karakteristikama i drugim odstupanjima od norme. Postoje muškarci s dodatnim Y kromosomom (XYY); Visoki su, agresivni i često imaju abnormalno ponašanje. Na Y kromosomu je identificirano nekoliko gena koji su odgovorni za regulaciju sinteze specifičnih enzima i hormona, a poremećaji u njima dovode do patologija spolnog razvoja. Postoji niz morfoloških znakova za koje se vjeruje da su određeni genima na Y kromosomu; među njima je i razvoj dlaka u uhu. Obilježja ove vrste prenose se samo muškom linijom: od oca do sina.
Genetska determinacija spola, određen skupom spolnih kromosoma, podržava jednaku reprodukciju žena i muškaraca. Doista, ženska jajašca sadrže samo kromosom X, jer žene imaju genotip XX na svojim spolnim kromosomima. Genotip muškaraca je XY, pa je rođenje djevojčice ili dječaka u svakom konkretnom slučaju određeno nosi li spermij X ili Y kromosom. Budući da tijekom procesa mejoze kromosomi imaju jednake šanse ući u gametu, polovica gameta koje proizvode muške jedinke sadrži X kromosom, a polovica Y kromosom. Stoga se očekuje da polovica potomaka bude jednog, a polovica drugog spola.
Treba naglasiti da je nemoguće unaprijed predvidjeti rođenje dječaka ili djevojčice, jer je nemoguće predvidjeti koja će muška spolna stanica sudjelovati u oplodnji jajne stanice: nositeljica X ili Y kromosoma. Stoga je prisutnost više ili manje dječaka u obitelji stvar slučajnosti.
Zakon neovisne raspodjele svojstava (treći Mendelov zakon) je povrijeđen ako se geni koji određuju različita svojstva nalaze na istom kromosomu. Takvi geni se obično nasljeđuju zajedno, tj. okovano nasljedstvo. Fenomen vezanog nasljeđivanja proučavali su Thomas Morgan i njegovi suradnici te se stoga naziva Morganov zakon.
Zakon T. Morgana može se formulirati na sljedeći način: geni koji se nalaze na istom kromosomu čine vezujuću skupinu i često se nasljeđuju zajedno, dok učestalost zajedničkog nasljeđivanja ovisi o udaljenosti između gena (što bliže, to češće).
Razlog poremećaja vezanog nasljeđivanja je crossing over koji se događa u mejozi tijekom konjugacije kromosoma. U tom slučaju homologni kromosomi izmjenjuju svoje dijelove, pa tako prethodno povezani geni mogu završiti na različitim homolognim kromosomima, što određuje neovisnu raspodjelu svojstava.
Na primjer, gen A vezan je za gen B (AB), a homologni kromosom sadrži recesivne alele odgovarajućih gena (ab). Ako tijekom procesa crossing overa homologni kromosomi gotovo nikada ne izmijene dijelove tako da jedan gen prijeđe u drugi kromosom, a drugi ostane u istom, tada takav organizam stvara gamete samo dvije vrste: AB (50%) i ab (50%). Ako dođe do izmjene odgovarajućih dijelova, tada će određeni postotak gameta sadržavati gene Ab i aB. Obično je njihov postotak manji nego kod neovisne raspodjele gena (kada su A i B na različitim kromosomima). Ako će kod neovisne raspodjele svih vrsta gameta (AB, ab, Ab, aB) biti po 25%, tada će u slučaju vezanog nasljeđivanja gameta Ab i aB biti manje. Što ih je manje, to su geni bliže jedan drugome na kromosomu.
Spolno vezano nasljeđivanje posebno se ističe kada se gen koji se proučava nalazi na spolnom (obično X) kromosomu. U ovom slučaju proučava se nasljeđe jedne osobine, a druga je spol. Ako je nasljedna osobina vezana za spol, tada se različito nasljeđuje tijekom recipročnih križanja (kada osobinu prvo posjeduje ženski roditelj, a zatim muški).
Ako majka ima genotip aa, a otac pokazuje dominantnu osobinu (definitivno postoji jedan gen A), tada će u slučaju spolne povezanosti sve kćeri imati dominantnu osobinu (u svakom slučaju, one će dobiti njegov jedini X kromosom od oca, a svi sinovi će imati recesivni) (od oca se dobiva Y kromosom koji ne sadrži odgovarajući gen, a od majke u svakom slučaju gen a.) Da osobina nije spolno vezane, onda bi među oba spola djece mogli biti vlasnici dominantne osobine.
Kada su geni koji se proučavaju povezani u autosomu, takvo se povezivanje naziva autosomnim. Povezivanje se naziva potpunim ako se roditeljske kombinacije alela ne prekidaju iz generacije u generaciju. To se događa vrlo rijetko. Obično se uočava nepotpuno vezano nasljeđivanje, koje krši i Mendelov treći zakon i Morganov zakon (u skraćenom obliku: geni koji se nalaze na istom kromosomu nasljeđuju se zajedno).
Geni na kromosomu raspoređeni su linearno. Udaljenost između njih mjeri se u centimorganima (cm). 1 cm odgovara prisutnosti 1% crossover gameta. Provodeći različita križanja i statističku analizu potomaka, znanstvenici identificiraju povezane gene, kao i udaljenost između njih. Na temelju dobivenih podataka izrađuju se genetske karte koje odražavaju lokalizaciju gena na kromosomima.
Vezano nasljeđivanje je pojava koreliranog nasljeđivanja određenih stanja gena koji se nalaze na istom kromosomu.
Ne postoji potpuna korelacija zbog mejotičkog crossing overa, jer se povezani geni mogu raspršiti u različite gamete. Crossing over se promatra u obliku odvajanja u potomstvu onih genskih alela i, sukladno tome, stanja svojstava koja su bila povezana kod roditelja.
Promatranja Thomasa Morgana pokazala su da vjerojatnost križanja između različitih parova gena varira, pa je došlo do ideje da se naprave mape gena na temelju učestalosti križanja između različitih gena. Prvu gensku mapu konstruirao je Morganov student, Alfred Sturtevant, 1913. godine koristeći materijal iz Drosophile melanogaster.
Udaljenost između gena koji se nalaze na istom kromosomu određena je postotkom crossing overa između njih i izravno je proporcionalna tome. Jedinica udaljenosti je 1% crossing overa (1 morganid ili 1 centimorganid). Što su geni udaljeniji jedan od drugoga na kromosomu, to će se križanje između njih češće događati. Maksimalna udaljenost između gena koji se nalaze na istom kromosomu može biti 49 centimorganida.
Povezane značajke
Vezane osobine su osobine koje kontroliraju geni smješteni na istom kromosomu. Naravno, oni se prenose zajedno u slučajevima potpune sprege.
Morganov zakon
Vezani geni lokalizirani na istom kromosomu nasljeđuju se zajedno i ne pokazuju neovisnu distribuciju
Prelazak preko
Međutim, homologni kromosomi mogu prelaziti (crossing over ili križanje) i razmjenjivati homologne regije. U ovom slučaju, geni jednog kromosoma prenose se u drugi, homologni njemu. Što su geni bliže jedan drugome na kromosomu, to je veza među njima jača i rjeđe dolazi do njihove divergencije tijekom crossing overa, i obrnuto, što su geni udaljeniji jedan od drugoga, to je veza između njih i slabija. to je češće moguć njegov poremećaj.
Broj različitih vrsta gameta ovisit će o učestalosti križanja ili udaljenosti između analiziranih gena. Udaljenost između gena izračunava se u morganidima: jedna jedinica udaljenosti između gena koji se nalaze na istom kromosomu odgovara 1% crossing overa. Ovaj odnos između udaljenosti i učestalosti križanja može se pratiti samo do 50 morganida.
27. Kromosomska teorija nasljeđa.
Kromosomska teorija nasljeđa je teorija prema kojoj su kromosomi sadržani u staničnoj jezgri nositelji gena i predstavljaju materijalnu osnovu nasljeđa, odnosno da je kontinuitet svojstava organizama u nizu generacija određen kontinuitetom njihove kromosome. Kromosomska teorija nasljeđivanja nastala je početkom 20. stoljeća. temelji se na staničnoj teoriji i koristio se za proučavanje nasljednih svojstava organizama putem hibridološke analize.
Utemeljitelj teorije kromosoma, Thomas Gent Morgan, američki genetičar, nobelovac. Morgan i njegovi studenti otkrili su da:
– svaki gen ima određeni lokus (mjesto) na kromosomu;
– geni na kromosomu nalaze se u određenom nizu;
– najbliže smješteni geni na jednom kromosomu su povezani, pa se nasljeđuju pretežno zajedno;
– skupine gena smještene na istom kromosomu tvore skupine veza;
– broj veznih skupina jednak je haploidnom skupu kromosoma u homogametnih jedinki i n+1 u heterogametnih jedinki;
– između homolognih kromosoma može doći do izmjene dijelova (crossing over); kao rezultat crossing overa nastaju gamete čiji kromosomi sadrže nove kombinacije gena;
– učestalost (u%) križanja između nealelnih gena proporcionalna je udaljenosti među njima;
– skup kromosoma u stanicama određenog tipa (kariotip) karakteristična je značajka vrste;
– učestalost križanja između homolognih kromosoma ovisi o udaljenosti između gena lokaliziranih na istom kromosomu. Što je ta udaljenost veća, to je veća frekvencija prijelaza. Za jedinicu udaljenosti između gena uzima se 1 morganida (1% crossing overa) ili postotak pojavljivanja crossover jedinki. Ako je ta vrijednost 10 morganida, može se reći da je učestalost križanja kromosoma na mjestima ovih gena 10% i da će nove genetske kombinacije biti identificirane u 10% potomaka.
Kako bi se razjasnila priroda položaja gena na kromosomima i odredila učestalost njihovog križanja, izrađuju se genetske karte. Karta odražava redoslijed gena na kromosomu i udaljenost između gena na istom kromosomu. Ovi zaključci Morgana i njegovih kolega nazvani su kromosomskom teorijom nasljeđa. Najvažnije posljedice ove teorije su moderne ideje o genu kao funkcionalnoj jedinici nasljeđa, njegovoj djeljivosti i sposobnosti interakcije s drugim genima.
Formiranje teorije kromosoma olakšano je podacima dobivenim proučavanjem genetike spola, kada su utvrđene razlike u skupu kromosoma u organizmima različitih spolova.
U ovoj lekciji ćemo pogledati nekoliko gena čije nasljeđivanje nije u skladu s Mendelovim zakonima, naučiti što je crossing over i vezano nasljeđivanje gena i razgovarati o njihovim značajkama.
Formiranje ovih gameta je ravnomjerno, što je predstavljalo osnovu za otkriće trećeg for-ko-ona Mende -la - za ne-zna-si-moje cijepanje znakova. Međutim, tijekom brojnih eksperimenata znanstvenici su saznali da postoje takvi parovi znakova, u kojima se dvije vrste gameta nalaze češće od druge dvije vrste gameta.
Pogledajmo to na konkretnom primjeru (slika 2).

Riža. 2. Drosophila muha ()
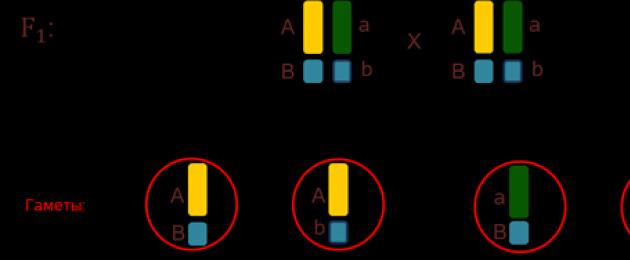
Kod malih vinskih mušica, dro-zo-phil - from-the-love-object-of-ge-ne-ti-kov - gen A from-ve-cha-et za sivo tijelo, recesivni gen A - za crnu boju tijela, dominantni gen B - za razvoj dugih krila, a recesivni jak gen in - za nerazvijena krila, odnosno krila ostaju u istom stanju.
Kod križanja dva ge-te-ro-zy-gota, koji imaju sivo tijelo, duga krila i tipove gena AaBb, u prvom redu generacije dobivaju tri tijela sa sivim tijelom i dugim krilima, kao i jedan organizam sa crnim tijelo i -cha-precizna krila (sl. 3).

Riža. 3. Križanje heterozigota AaBb ()
Ubuduće obratite pozornost na činjenicu da su znakovi u or-ga-dnou značajniji bol -sher od kro-mo-soma, u kojem su ti znakovi lo-ka-li-zo-va-ny. Zaključili su to, očito, u jednom kromosom postoji veliki broj gena. Geni smješteni u jednom chro-mo-so-me čine jednu grupu - kvačilo, a zatim najčešće zajedno (sl. 4).

Riža. 4. Vezani geni ()
Ovaj je obrazac uspostavio američki gene-ne-ti-k To-ma-s Gen-Mor-ga-n u na-cha- le 20-ih posljednje stoljeće. Prema tome, ako geni leže u istom kromosomu, to znači da će se nasljeđivati zajedno (slika 5).

Riža. 5. Lančano nasljeđivanje ()
D - gen koji označava sivo tijelo; d - crno tijelo; F - duga krila; f - za-precizna krila.
Kao rezultat takvog križanja imamo tri or-ga-niz-ma, sivog tijela i dugih novih krila, i jedan or-ga-izam, crnog tijela i vrlo preciznih krila.
Ge-ne-ti-ki obratite pozornost na činjenicu da se u potomcima takvih muha još uvijek pojavljuju or-ga-dna sa sivim tijelom i iza-preciznih krila, kao i s crnim tijelom i dugim krilima. Objašnjenje za to pronađeno je tijekom proučavanja razvoja stanica - taj se proces naziva "mejoza".
U profazi 1 postoji fenomen konjugacije ili konvergencije kromosoma, za što može slijediti križ-grijeh-go-ver- izmjena dijelova homolognih kromosoma (slika 6).

Riža. 6. Cross-syn-go-vera proces ()
U re-zul-ta-th e-ra-zu-yut-sya cross-so-faithful ga-me-you. Or-ga-low-we, koji su uključeni u rezultat fuzije takvih cross-sovere gameta, nazivaju se "re-com-bi-nant-nye or-ga-niz-we." Budući da se cross-syn-go-ver ne događa nakon svake konjugacije, tada je broj cross-syn-go-ver-gameta značajan, ali manji od broja necros-verificiranih gameta, omjer je oko 20 prema 80 %.
Tijekom ex-peri-men-ta To-ma-su Gent Mor-ga-nu uspio je dokazati da često cross-sin-go-ve-ra između gena -mi izravno o udaljenosti između njih u kromosom, to jest, možemo reći da što su geni kromosomski udaljeniji jedan od drugoga, to se među njima češće javlja unakrsna sin-go-ver.
Ovo je otkriće pos-vo-li-lo la-bo-ra-to-rii To-ma-sa Mor-ga-na za razvoj metode, pos-la-yu-schy za izgradnju hro-mo-nekih karata , odnosno označavaju mjesto gena u kro-mo-somu za različite organizme -niz-mov.
Chro-mo-some karte stvorene su praktički za sve poljoprivredno važne životinje i biljke, rad na ovom desnom dijelu i dalje se nastavlja, iako ge-ne-ti-ki već koristi ne samo me-dom od gi -bri- di-za-cije, ali im i drugi s vremena na vrijeme imaju pristup.
Bibliografija
- Mamontov S.G., Zakharov V.B., Agafonova I.B., Sonin N.I. Biologija. Opći obrasci. - Droplja, 2009.
- Ponomareva I.N., Kornilova O.A., Chernova N.M. Osnove opće biologije. 9. razred: Udžbenik za učenike 9. razreda općeobrazovnih ustanova / Ured. prof. U. Ponomareva. - 2. izdanje, revidirano. - M.: Ventana-Graf, 2005
- Pasechnik V.V., Kamensky A.A., Kriksunov E.A. Biologija. Uvod u opću biologiju i ekologiju: Udžbenik za 9. razred, 3. izd., stereotip. - M.: Bustard, 2002.
- Polnaja-jenciklopedija.ru ().
- Jaslice.me().
- Biorepet-ufa.ru ().
Domaća zadaća
- Koji se parovi gena ne pokoravaju Mendelovim zakonima?
- Kakav je uzorak gena koji leže na istom kromosomu?
- Što je crossing over?
- U kontaktu s 0
- Google+ 0
- u redu 0
- Facebook 0






