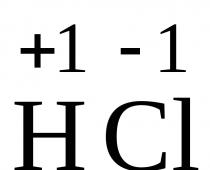Se även "Mesoderm" i andra ordböcker
(mesoderma, LNE; meso- + grekisk dermahud; synonym mesoblast) - det mellersta groddskiktet som bildas hos däggdjur genom tillväxten av den primära strimmen i form av ett lager av celler mellan ekto- och endoderm.
extraembryonala mesoderm (m. extraembryonicum, LNE) - en del av mesodermen, en del av embryots temporära (provisoriska) organ - de embryonala membranen och vitelline
splanchnic mesoderm (m. splanchnicum, m. viscerale, LNE; synonym: splanchnomesoderm, splanchnopleura) - del av den laterala mesodermen, från vilken de viscerala skikten av lungsäcken, bukhinnan och mesenteriet, hjärtat, endotelet av blodkärl, binde- och glatt muskelvävnad av inre organ bildas.
dermal mesoderm (m. paraxiale, LNE; m. dermale) - del av mesoderm, som därefter bildar bindvävsdelen av huden (dermis).
dorsal mesoderm (m. dorsale; syn. M. parachordal) - en del av mesoderm, som är parat förtjockningar på båda sidor av notokorden, bildar somiter.
larv mesoderm (m. larvale; lat. larvmask...
(från meso... och dermis), mesoblast, mellangroddlager hos flercelliga djur (förutom svampar och coelenterater). Ligger mellan ektodermen och endodermen. Olika grupper av djur utvecklas olika sätt (se Gastrulation). Hos plattmaskar och nemerteaner ger M:s ränder bindväv som fyller ut utrymmet mellan de inre delarna. organ, hos annelider och de flesta andra ryggradslösa djur är M.-ränderna uppdelade i parade somiter med en sekundär kavitet - coelom. Hos ryggradsdjur, under neurulationsperioden, från sidorna av notochord primordium, är M- uppdelad i dorsala (primära) segment - somiter, nefrotomer och osegmenterad buk M. - laterala plattor. Mellan de två bladen av var och en av dem bildas en coelom. M. och dess derivat har en inducerande effekt ((se Induktion) på utvecklingen av derivat av ektodermen och endodermen och upplever i sin tur ett inducerande inflytande från deras sida (se Germ Layers).
embryots mellersta lager från vilket många kroppsvävnader utvecklas.
(Källa: "Ordbok över främmande ord som ingår i det ryska språket." Pavlenkov F., 1907)
1. Typ av groddvävnad.
(mesoderm) - det mellersta groddskiktet av ett embryo i de tidiga utvecklingsstadierna. Det fungerar som en källa till utveckling av brosk, muskler, ben, blod, njurar, könskörtlar och deras kanaler och bindväv. Mesodermen är uppdelad i två lager: den yttre somatiska och djupa, viscerala, åtskilda av en hålighet - coelom (coelom), som blir kroppshålan. Den dorsala somatiska mesodermen segmenteras i ett antal somiter. se även Mesenchyme. - Mesodermal (mesodermat).
MESODERM
(Förlåt... Och dermis)(mesoblast), det mellersta groddskiktet hos flercelliga djur (förutom svampar och coelenterater) och människor. Från M. utvecklas muskler, brosk, skelett, organ för blod och lymfbildning, sekret, könsorgan etc. Ektoderm, Endoderm.
MESODERM
MESODERM, det mellersta GERMINAL-skiktet av vävnad som bildas i det tidiga utvecklingsstadiet av ett befruktat ägg (OVA) i nästan alla flercelliga organismer. I senare utvecklingsstadier ger det upphov till muskler, blod och bindväv. De andra groddskikten är ECTODERM och ENDODERM.
mesoderm
mesoblast
Ordbok för ryska synonymerMesoderm
mesod\"erma, -s
Rysk stavningsordbok. / Ryska vetenskapsakademin. Institutet rus. språk dem. V. V. Vinogradova. - M.: "Azbukovnik". V. V. Lopatin (exekutiv redaktör), B. Z. Bukchina, N. A. Eskova och andra.. 1999 .
MESODERM (från meso... och dermis) (mesoblast) - det mellersta groddskiktet hos flercelliga djur (förutom svampar och coelenterater) och människor. Från mesoderm muskler, brosk, ben, organ av blod och lymfbildning, sekret, könsorgan, etc. utvecklas. Ektoderm, Endoderm.
Mesoderm
mesode e/ rma, -s
Tillsammans. Isär. Avstavat.. B. Z. Bookchina.
Mesoderm
(mesoderma, LNE; meso- + grekisk dermahud; synonym mesoblast) mellangroddlager, bildat hos däggdjur genom tillväxten av den primitiva strimman i form av ett lager av celler mellan ekto- och endoderm.
Mesoderm se bakterieblad.
Mesoderm (mesoderma, LNE; meso- + grekisk dermahud; synonym mesoblast)
det mellersta groddskiktet som bildas hos däggdjur genom tillväxten av den primitiva strimman i form av ett skikt av celler mellan ekto- och endoderm. Extraembryonala mesoderm(m. extraembryonicum, LNE) - en del av M., en del av embryots temporära (provisoriska) organ - de embryonala hinnorna och gulesäcken. Intern mesoderm(m. splanchnicum, m. viscerale, LNE; synonym: splanchnomesoderm, splanchpopleura) - en del av den laterala M., från vilken de viscerala skikten av lungsäcken, bukhinnan och mesenteriet, hjärtat, endotelet av blodkärlen, bindande och släta muskelvävnad av inre organ bildas. Dermal mesoderm (... Mesoderm (från Meso... och grekiskt derma - hud)
mesoblast, mellangroddlager (Se Germlager) hos flercelliga djur (förutom svampar och coelenterater) och människor. Som ett resultat av gastrulation (Se Gastrulation) ligger den mellan det yttre groddskiktet - ektodermen (Se Ektoderm) och den inre - endodermen (Se Endoderm). Hos protostomer (Se Protostomes) djur (de flesta ryggradslösa djur) bildas muskeln genom den teloblastiska metoden - från stora celler - teloblaster, som ligger mellan ektodermen och endodermen vid embryots bakre ände och kommer in i den primära kroppshålan under gastrulation, där de förökar sig och förvandlas till två mesodermala ränder. Hos de flesta deuterostomes (se Deuterostomes) djur - tagghudingar, brachiopoder ...
Groddlager, eller groddlager - lager av embryots kropp hos flercelliga djur, som bildas i processen och ger upphov till olika organ och vävnader.
De bildas i processen för differentiering av liknande homogena celler
Gastrulation- utbildningsprocess två groddlager(ento- och ektoderm).
Under gastrulation rör sig alla celler och bildas gastrula- tvålagers embryosäck, inuti vilken det finns en hålighet - gastrocel, ansluten med den primära munnen ( blastopore) med den yttre miljön.
Gastrulation slutar med bildandet av det tredje groddskiktet - mesoderm, belägen mellan ekto- och endoderm.
I de flesta organismer (förutom coelenterater) bildas tre groddlager:
- extern - ektoderm,
- internt - endoderm Och
- genomsnitt - mesoderm.
Efter avslutad gastrulation bildar embryot ett komplex av axiella organ: neuralrör, notokord och tarmrör. Det här är scenen neurulae.
Utbildning groddlager- början på omvandlingen av en flercellig organism till en organism där cellerna blir differentierade och i framtiden bildar vävnader och organ.
Så först börjar zygoten dela sig, vilket ökar antalet celler. Efter att ha fått tillräcklig massa börjar kroppen nästa steg - cellerna börjar röra sig - de flyttar till periferin och bildar blastodermisk vesikel.
Vid ena kanten av denna vesikel grupperas celler och bildar en inre hålighet - det är inre groddlager - endoderm.
Embryots yttre celler (yttersta lagret) - ektoderm.
Lagret av celler mellan dessa två groddlager är mesoderm, dessa celler bildas dels från ekto-, dels från endoderm.
- Denna uppdelning av löv är typisk för alla högre djur;
- på enkla djur- endast y och - 2 groddlager(extern och intern).
Här är en exempelfråga från Unified State Exam i biologi bara på ämnet:

1. från ektodermen bildas: örat och hjärnan;
2. från endodermen - lever, lungor, tarmar, mage, bukspottkörteln;
3. från mesodermen - muskler, blodkärl, ben.
Groddlager beskrevs först i en rysk akademikers arbete X. Panderaår 1817, som studerade kycklingembryots embryonala utveckling. En särskilt viktig roll i studiet av ryggradsdjurens groddlager spelades av de klassiska verken av en annan rysk akademiker - Carla Bara, som visade att groddlager också finns i embryon från andra ryggradsdjur (fiskar, amfibier, reptiler).
Ontogenes av någon organism kännetecknas av bildandet av groddlager. Hos primitiva organismer som coelenterater och svampar består embryot av endast två lager: endoderm och ektoderm. Med tiden utvecklar mer progressiva former av organismer ett tredje lager - mesodermen.
Vad är mesoderm?
Ontogenes är den sekventiella utvecklingen av embryot, som åtföljs av ett antal förändringar i morfologin och anatomin hos den framtida unga organismen. Mesoderm är ett groddlager som spelar en viktig roll i bildandet av många organ och vävnader. Det är inte för inte att sådana primitiva flercelliga djur som hydra, maneter, koraller eller svampar kallas tvåskiktsdjur, eftersom de under ontogenesprocessen endast bildade två groddlager.
Mesoderm bildning
Processen för initiering av det mellersta groddskiktet skiljer sig mellan olika taxonomiska grupper. Det finns tre mest välkända sätt på vilka mesoderm bildas: teloblastisk, enterocoelös och ektodermal.
1. Den teloblastiska vägen för mesodermutveckling är karakteristisk för många protostomer och är baserad på bildandet av blastomerer. Några av dem är specialiserade på att lägga det mellersta groddskiktet, som i slutändan tar formen av två längsgående parallella band. Dessa band ger upphov till mesoderm.
2. Den enterocelösa metoden är fundamentalt annorlunda genom att mesodermprekursorcellerna bildar en invagination (invagination) tillsammans med endodermen. Denna invagination i framtiden bildar den primära tarmen. Gränsen mellan de två skikten förblir oskiljbar under lång tid, och först efter en lång tidsperiod kopplas mesodermen som ett oberoende skikt från endodermen. Denna utvecklingsmetod är karakteristisk för djur som lansett eller sjöstjärna.
3. Den ektodermala metoden för mesodermutveckling är karakteristisk för sådana typer av djur som reptiler, fåglar och däggdjur (inklusive människor). Summan av kardemumman är att efter invagination bildas endast endoderm. Om du föreställer dig ett tvärsnitt av embryot, kommer fritt utrymme att uppstå mellan endo- och ektoderm efter gastrulation (bildning av invagination). Celler av ektodermalt ursprung "knoppar" där, vilket ger upphov till ett nytt groddskikt.

Morfologi av mesoderm
Mesodermen spelar en stor roll i bildandet av embryot. Inom biologin är detta ett bra evolutionstecken, eftersom skillnaden i morfologin hos det mellersta groddskiktet i olika används i taxonomi.
Om vi betraktar två längsgående band som bildas under det teloblastiska utvecklingsläget, kommer mesodermen att representeras av metameriskt upprepade sektioner. Den dorsala sidan av varje sådant band är uppdelad i somiter, den laterala sidan i nefrotomer och den ventrala sidan i splanknotomer.

Vilken roll spelar mesoderm? Mänskliga organ bildade av mesoderm
Varje groddlager är ett slags föregångare till den framtida organismens organsystem och vävnader. Topologin hos de bildande bladen bestämmer till stor del deras framtida öde. Eftersom mesoderm är det mellersta germinala lagret, deltar det i bildandet av vävnader och organ som är belägna mellan integumentet hos en person och de innersta lagren av kroppen. Vilka strukturer är av mesodermalt ursprung?

Slutsats
Mesodermen är ett komplext embryo som i slutändan ger upphov till många vitala organ och vävnader. Mellanbladets bildande och utveckling skiljer sig åt hos olika djur, och detta är en av de evolutionära egenskaperna. Närvaron av mesoderm indikerar att djuret är treskiktigt, vilket är ett signifikant tecken på gruppens framsteg.
MESODERM(från grekiska mesos - mellersta och derma - hud), mellersta groddskiktet (se. groddlager), I klassisk mening är det en samling celler som bildar väggen i den sekundära kroppshåligheten, eller coelom, i embryot. Från sammansättningen av M. i de tidiga utvecklingsstadierna, rudimentet av den s.k. muskuloskeletala systemet, det vill säga all bindväv, inklusive blod och lymfa. Denna bakterie, som kallas mesenkym(se), sticker ut i form av isolerade celler eller separata cellkomplex. I vissa fall kan den inte skarpt avgränsas från det övriga M., som har b. inklusive naturen hos kontinuerliga cellskikt - M.-bildning hos ryggradsdjur spårades först av Kolliker (Kolliker, 1879) i embryot hos kycklingen och däggdjuren. Den exakta formuleringen av själva konceptet i förhållande till alla klasser av ryggradsdjur och många ryggradslösa djur gavs av Hertwigs (O. och R. Hertwig, 1881) baserat på den jämförande embryologiska metoden. M. hos ryggradsdjur bildas i de flesta fall i form av ett bilateralt symmetriskt rudiment längs kanterna blastopore(se) eller från motsvarande område enligt teorin om blastopore (Hertwig) (marginal hack av selachier och benfiskar, primär platta av reptiler, primär strimma av fåglar och däggdjur). Detta sk den peristomala mesodermen växer sedan i sidled och framåt på sidorna av ryggsträngsrudimentet, som bildas framför blastoporen och ingår i vissa utvecklingsstadier i primärtarmens vägg (cefalisk process i fostervatten); sista delen av M. är den s.k. parachordal, eller g och -stral M. [se. separat bord (art. 743-744), fig. 1]. Endast i lansetten, liksom hos vissa ryggradslösa djur, isoleras m. efter tillslutningen av blastoporen i form av segmenterade parade utsprång från primärtarmens vägg. I alla andra fall har den embryonala grodden hos ryggradsdjur initialt karaktären av en kontinuerlig parad cellplatta, inkilad mellan embryots ekto- och endoderm. Kroppshålan uppträder senare genom att M.:s cellskikt delas i två blad.. M.:s rudiment, som växer allt längre i båda riktningarna från ursprungsplatsen, är uppdelad i flera sektioner som har olika prospektiv betydelse, d.v.s. att därefter bilda olika differentieringsprodukter. Den på båda sidor om notokorden och neuralröret liggande ryggdelen av M. genomgår metamerisk uppdelning (segmenterad) i s.k. dorsala segment, eller m i o till m s. Segmenteringen börjar från embryots huvudände och sprider sig gradvis till den kaudala änden, och antalet segmentpar ökar gradvis med embryots ålder. Myotomer finns kvar en tid i anslutning till myotomens icke-segmenterade ventrala sektioner, splanknotomer och, genom segmenterade mellanliggande avsmalnande områden, de s.k. segmentben eller nefrotomer. I myotomerna bör man också urskilja ett speciellt medio-ventralt område, sklerotomen, som då sticker ut från själva muskelns sammansättning och är en av källorna till bildning. mesenkym(centimeter.). Endast i lansletten bildas icke-segmenterade splanknotomer genom sekundär sammansmältning av de primära segmentens ventrala sektioner. Den sekundära kroppshålan, eller coelom, når sin största utveckling i splanknotomer, i vilka den kallas splanchnocoel. Den del av splanknotomen som gränsar till ektodermen, som delas som ett resultat av bildandet av coelom i två lager, kallas parietal (parietal), trunk, hudfiber-lgst eller somatopleura. Bladet som gränsar till endodermen kallas det viscerala (inre), tarmfiberbladet eller splanchnopleura. Det bör noteras att det mesta av myotomväggen, vänd mot ektodermen och betecknad som hudplattan, liksom den ovan nämnda sklerotomen, representerar den andra källan till mesenkymbildning och separeras snart från själva mesenkymets sammansättning, resten av skäret kallas muskellagret och NK och . De parietala och viscerala skikten av splanknotomerna innehåller också diffusa mesenkymala rudiment, som gradvis separeras från sin sammansättning. Alla dessa släktskap är tydligast iakttagna i embryon av selachianer, medan de hos andra ryggradsdjur ofta till stor del är skymd och kan endast tydas genom jämförelse med dessa senare. Hos ryggradsdjur, som under embryonalt liv har speciella hjälporgan - äggulablåsan och embryonala membran (se Germ),-ventrala sektioner av splanknotomer är en del av dessa senare och kallas extraembryonala mesoderm. Detta senare kan i sin struktur och delvis i sitt vidare öde inte skarpt skiljas från mesenkym(centimeter.). Hos människor bildas extraembryonala mesenkym (eller mesenkym) i mycket tidiga utvecklingsstadier, oavsett dess embryonala del, och uppvisar uppenbarligen inte bilateral symmetri.Den del av kroppshålan som begränsas av den extraembryonala mesodermen betecknas som den extraembryonala coelomen. Tack vare de senaste framgångarna inom experimentell embryologi måste vi erkänna (Y. Spomann, O. Mangold, W. Vogt, L. Graper, etc.) att det cellulära materialet som M. borde bildas redan är på stadium blastula(se) eller i början av gastrulationsprocessen (se. Gaetrula) har strikt lokalisering. Detta cellulära material kallas presumtiv mesoderm. I lansetten, enligt E. Conclin, är det även i en befruktad äggcell möjligt att urskilja en speciell sektion av protoplasman, från vilken sedan bildas M. Med ytterligare differentiering från olika delar av den embryonala M. utvecklas mycket olika vävnader. hos ryggradsdjur. Myotomer ger upphov till frivilliga somatiska muskler, nefrotomer, eller segmentella ben, ger upphov till epitelet av utsöndringsorganen och delar av hanens utsöndringskanaler och hos vissa lägre ryggradsdjur den kvinnliga reproduktionsapparaten. splanknotomerna (se. mesotel) serösa kaviteter, sk. interrenala organ (se Binjurarna), hjärtmuskeln och hos högre ryggradsdjur, tydligen delar Müllers traktat(s "m.) och dess derivator. Därför kan man inte låta bli att erkänna att M. i huvudsak är ett kombinerat begrepp som innehåller mycket heterogena rudiment. Detta ger upphov till att många biologer inte anser det möjligt att titta på M. som i ett enda groddlager. Belyst.: CClin E., The embryology of Amphio-xus, Journ. av morf., v. LIV, 1932; G r a p e r L., Die Primitiventwicklung des Htihnchens nach stereokinemato-graphischen Untersuchungen, kontrolliert (lurch vitale Farbmarkierung und verglichen mit der Entwicklung ander Wirbeltiere, Arch. f. Entwicklungsmech. d. Org., B. CXVI, u. Hcrtwig O. 1929; B,., Die Colomtheorie, Versuch einer Erkbirung des mittleren Keimblattes, Jenaer Zeitschr. f. Naturw., B. XV, 1881; de även Die Lehre von den Keimbliiltern (Hndb. d. vergleichenden u. experi-mentellen Untwieklungierehre der Wirbeltierehre , hrsg. v. O. Hertwig B I, T. 1, Jena, 1906); E, a b 1 C, Theorie des Mesoderms, Morphol. Jahrb., B. XV u. XIX, 1889-93; V o g t W. , Gestaltungsanalyse am Amphibienkeim mit ortlicher Vitalfarbung, Arch.f.Entwicklungsmech.d.Org., B.CXXX, 1929.H.Hos alla djur, med undantag för coelenterater, i samband med gastrulation (parallellt med den eller i nästa stadium orsakad av gastrulation), uppträder ett tredje groddlager, mesodermen. Detta är en uppsättning cellulära element som ligger mellan ektodermen och endodermen, det vill säga i blastocoele. Således blir embryot inte tvålager, utan trelager. Hos högre ryggradsdjur uppträder en treskiktad struktur av embryon redan under gastrulationsprocessen, medan i lägre chordater och alla andra typer, som ett resultat av gastrulation, bildas ett tvåskikts embryo.
Frågor om vägarna för mesodermbildning hos olika djur har länge varit intressanta för både jämförande anatomer och embryologer. I allmänhet kan de anses lösta, dock inte i termer av orsakerna till motsvarande morfogenetiska processer, utan i planet för den formella morfologiska beskrivningen av dessa processer. Om vi abstraherar från alla de olika detaljerna i bildandet av mesoderm i olika djur, kan vi fastställa två fundamentalt olika sätt för dess uppkomst: teloblastisk, dess egen -™™“ och Protostomia, och enterocoelous, karakteristisk för Deutes a. i protostomer, under gastrulation, vid gränsen mellan ektoderm och endoderm, på sidorna av blastoporen, finns det redan två stora celler (eller flera sådana kroppsceller)
Ris. 51. Grovt diagram över bildningen av mesoderm i protostomer (A) och deuterostomer (B) (enligt V.M. Shimkevich, 1925, modifierad):
/ - ektoderm, 2 - mesenkym, 3 - endoderm, 4 - teloblast (L) och coelomisk mesoderm (5)
sprängningar), separerar små celler från sig själva (på grund av delningar) (Fig. 51, L, Fig. 69). Således bildas mittskiktet - mesodermen. Teloblaster, som ger upphov till nya generationer av mesodermceller, stannar kvar i den bakre änden av embryot. Av denna anledning kallas denna metod för mesodermbildning teloblastisk (från grekiskans telos - ände).
Med enterocoel-metoden uppträder en uppsättning celler av den utvecklande mesodermen i form av fickliknande utsprång av primärtarmen (utskjutande av dess väggar in i blastocoel, Fig. 51, B, 4). Dessa utsprång, i vilka delar av den primära tarmhålan kommer in, separeras från tarmen och separeras från den i form av påsar. Kaviteten i säckarna förvandlas till en helhet, det vill säga till en sekundär kroppshålighet; coelomiska säckar kan delas in i segment.
Denna beskrivning av metoderna för ursprunget för det mellersta groddskiktet återspeglar inte hela variationen av variationer och avvikelser som är strikt naturliga för enskilda grupper av djur. Liknande den teloblastiska metoden, men endast externt, är metoden för bildning av mesoderm inte genom att dela teloblaster, utan genom uppkomsten vid kanterna av blastoporen av en oparad tät primordium (grupp av celler), som därefter delar sig i två symmetriska ränder av celler. Med enterocoel-metoden kan mesoderm rudiment vara parat eller oparat; i vissa fall bildas två symmetriska coelomic säckar, och i andra bildas först en gemensam coelomic säck, som sedan delas i två symmetriska halvor.
Det har redan sagts om de speciella utvecklingsprocesserna för nematoder och andra djur, i förhållande till vilka det skulle vara konstgjort att tillämpa begreppet "groddlager" - i dem, kringgå bildandet av cellulära groddlager, rudimenten av framtida organ är isolerade i form av separata blastomerer.
På grund av dess betydelse för embryologin i allmänhet och för att förstå processerna för organutveckling, kommer nästa kapitel att ge en jämförande embryologisk översikt över gastrulationsprocesserna hos olika djur, med lämpliga justeringar av de alltför förenklade klassiska idéerna om groddskikten, i speciellt om den enterocoelösa metoden för mesodermbildning.
Gastrulation är ett stadium i embryots utveckling. Groddskikten är inte något separat från varandra, deras uppkomst och ytterligare förändringar sker på grund av det ömsesidiga beroendet av embryots delar. Groddskikten, som samlingar av celler, skiljer sig från varandra inte bara i sin position i embryots allmänna system, utan också i vissa cytologiska egenskaper. Samtidigt övertygar experiment att deras öde fortfarande kan förändras, vilket tvingar dem att "bygga" cellulära system och organ som är ovanliga för dem (se kapitel XI och XVII).
Under den normala utvecklingen av embryon fortsätter groddlagren, som interagerar med varandra under påverkan av de integrerande influenserna från embryot som helhet, att differentiera sig i en viss riktning, och var och en av dem deltar i uppkomsten av rudimenten av vissa organ och organsystem. Vi kan prata om ett nytt stadium i utvecklingen av embryon - organogenes.
I hela djurriket kommer vissa organ från samma groddlager. Undantag från denna lag, som kommer att diskuteras senare, beror på förändringar i ontogenes, i samband med vissa unika vägar för djurens evolution. De anses vara homologa formationer i embryot. För groddlager, se kap. VII-IX.
Derivat av ektoderm. De flesta av cellerna som utgör den yttre piasten, multiplicerar och differentierar sig i enlighet därmed, förblir på ytan och deltar i utvecklingen av kroppens integument. Från dem bildas: yttre epitel, hudkörtlar, ytskikt av tänder, kåta fjäll. d. Nästan alltid utvecklas varje organ från de cellulära elementen i två, eller till och med alla tre bakterielagren. Till exempel utvecklas däggdjurshud från ekto- och mesoderm.
Som regel "sjunker" en stor del av den primära ektodermen (upp till en tredjedel eller mer av hela ytan av amfibieembryon), på grund av speciella morfogenetiska processer, inuti, under det yttre epitelet och ger upphov till hela nervsystemet systemet. Hos många djur invagineras ektodermen vid de främre och bakre ändarna av kroppen mot de främre och bakre ändarna av tarmen som utvecklas från endodermen (midgut). Dessa invaginationer bryter in i mellantarmens hålighet och bildar stomodeum (förtarm) och proctodeum (baktarm).
Endodermderivat. Det inre groddskiktet, som skiljer sig åt i samband med andra delar av embryot, utvecklas till epitelet i mellantarmen och dess matsmältningskörtlar. Utveckling av epitelet i andningssystemet (gren och
lungor) hos olika ryggradsdjur har inte spårats lika fullständigt och är ännu inte så tydlig i detalj. Det är obestridligt att detta epitel utvecklas från förtarmen. Det kan dock inte sägas kategoriskt att det är helt och hållet av endodermalt ursprung, eftersom
huru den prechordala plattans cellulära material utan tvivel deltar i dess uppkomst (se s. 126 etc.).
Mesodermderivat. Alla andra organ som inte tidigare listats utvecklas från mesodermen: all muskelvävnad, var de än befinner sig (kroppsväggen, tarmarna och andra formationer), alla typer av bindväv, brosk- och benvävnad, utsöndringsorganens kanaler, peritoneum i kroppshåla, cirkulationssystemet , en del av vävnaderna i äggstockarna och testiklarna. Med utvecklingen av motsvarande organ uppstår specifik differentiering av de cellulära elementen i mesodermen. Hos de flesta djur uppträder mittskiktet inte bara i form av en samling celler som bildar ett kompakt epitelliknande skikt, d.v.s. själva mesodermen, utan också i form av ett löst komplex av spridda, amöbaliknande celler. Denna del av mesodermen kallas mesenkym. Mesoderm och mesenkym skiljer sig från varandra i sitt ursprung, det finns ingen direkt koppling mellan dem, de är inte homologa. Mesenkym är mestadels av ektodermalt ursprung, medan mesoderm börjar med endoderm. Hos ryggradsdjur är dock en mindre del av mesenkymet av ektodermalt ursprung, medan huvuddelen av mesenkymet har ett gemensamt ursprung med resten av mesodermen. Hos många djur med spiralklyvning uppträder mesenkym under klyvningen. I tagghudingar är källan till mesenkym mikromerer och endoderm. Celler i botten av den utvecklande primära tarmen migrerar in i blastocoel.
Trots sitt annorlunda ursprung från mesoderm kan mesenkym betraktas som en del av mellanskiktet. Det spelar en stor roll i bildandet av larven och definitiva organ.
För att förstå de frågor som diskuteras ytterligare är det nödvändigt att ha en uppfattning om en viktig formation - coelom, kroppens sekundära hålighet. Hos alla djur som kännetecknas av en coelom ger de ihåliga coelomiska säckarna upphov till mesodermen. Det har redan sagts att med det enterocoeliska ursprunget av mesoderm, bildas coelomiska påsar genom att förändra, differentierande fickliknande utsprång i primärtarmen. I teloblastiska och liknande metoder, när mesodermala sladdar bildas, uppstår ett gap inuti dem, som så småningom förvandlas till en helhet. Coelomic påsar bildas symmetriskt på sidorna av tarmen. Väggen i varje coelomisk säck som vetter mot tarmen kallas splanchnopleura. Väggen som vetter mot embryots ektoderm kallas somatopleura.
Således, under utvecklingen av embryot, olika
Ris. 52. Schema för organogenes av embryon från högre ryggradsdjur (enligt K. Waddington; 1957):
/ - neuralrör, 2 - somit, 3 - notokord, 4 - tarm, 5 - lateral mesoderm, som helhet, 7 - epidermis, c - svalg, 9 - gälslitsar, 10 - optisk vesikel, 11 - hjärna
personliga håligheter som har viktig morfogenetisk eller funktionell betydelse. Först uppträder Baers hålighet, förvandlas till den primära kroppshålan - blastocoel, sedan, i samband med gastrulationsprocesserna, uppträder gastrocoel (eller maghålan), och slutligen, hos många djur - coelom. Med bildandet av gastrocoel och coelom blir blastocoel allt mindre, så att allt som återstår av den tidigare primära kroppshålan är luckor i utrymmena mellan väggarna i tarmen och coelom. Hos vissa djur förvandlas dessa slitsar till håligheter i cirkulationssystemet. Magsäcken förvandlas så småningom till mellantarmshålan.
Med enterocoel-metoden för att separera mesoderm och coelom på bekostnad av gastrocoel, uppträder dessutom en sekundär kroppshålighet.
Gastrulationsprocesserna leder direkt till en period av organogenes. Hos vissa djur bildas organ och organsystem, som gradvis får definitiv betydelse, medan hos andra djur först bildas organ som är karakteristiska för larven, därefter sker metamorfos (se kapitel X) och bildningsprocesserna för de definitiva organen i vuxen organism förekommer.
På grund av avsaknaden av en enhetlig plan i strukturen av embryona hos alla ryggradslösa djur är det omöjligt att ge ett diagram över ett abstrakt embryo av ett ryggradslöst djur. Ett diagram över det sena embryot av ett ryggradsdjur visas i fig. 52.
- I kontakt med 0
- Google+ 0
- OK 0
- Facebook 0