Определения терминов, приведенные в этом словаре, не всегда являются абсолютно точными и строгими с научной точки зрения, хотя для популярной книги точность вполне достаточная. Давать строгие определения биологическим понятиям - крайне неблагодарное дело. Как ни изворачивайся, в любом определении все равно найдутся неточности и исключения, слишком уж сложны и разнообразны объекты биологических исследований.
абиогенный (процесс) - не связанный с живыми организмами,осуществляемый без участия живого,
абиогенез - происхождение жизни из неживой материи в результате естественных процессов развития (химической эволюции).
автотрофы - организмы, умеющие превращать неорганический углерод в органические соединения, то есть осуществлять фиксацию углерода. Глава 2, стр. 95.
адаптация - приспособление. Термин используется в двух смыслах: 1. орган, признак, свойство, имеющее приспособительное значение: "Густой мех - адаптация к холодному климату"; 2. процесс приобретения таких признаков: "В ходе адаптации к холодному климату животное приобрело густой мех". Когда хотят подчеркнуть, что речь идет не о приспособительных признаках, а именно о процессе их приобретения, иногда пользуются термином "адаптациогенез".
аллель - вариант гена. В популяции каждый ген может существовать в виде множества разных аллелей. У гаплоидных организмов каждый ген присутствует в единственном экземпляре, то есть имеется только один аллель данного гена. У диплоидных организмов каждый ген присутствует в двух копиях. Если обе копии гена представляют собой один и тот же аллель, такой организм называют гомозиготным по данному гену. Если две копии различаются, то есть представляют собой разные аллели, такой организм называют гетерозиготным.
анаэробы - организмы, не нуждающиеся в кислороде и обитающие в бескислородных условиях.
антиген - чужеродное вещество, обычно белок или углевод, которое при введении в организм животного вызывает появление специфических антител, умеющих распознавать и связывать данный антиген.
антикодон - три нуклеотида (триплет) в молекуле транспортной РНК, комплементарные какому-либо кодону матричной РНК (см. "генетический код"). В ходе трансляции (синтеза белка) транспортные РНК, несущие определенные аминокислоты, поочередно присоединяются своими антикодонами к кодонам мРНК. Специальные ферменты прикрепляют очередную аминокислоту, принесенную молекулой тРНК, к синтезируемой молекуле белка. Так осуществляется считывание генетического кода, или перевод последовательности нуклеотидов мРНК в последовательность аминокислот молекулы белка. Глава 1, стр. 59, 61.
антитела - защитные белки, которые умеют узнавать определенных бактерий, вирусов, а также любые чужеродные белки и многие углеводы, и прикрепляться к ним, что приводит к обезвреживанию самих возбудителей или выделяемых ими токсинов. Молекулы, распознаваемые антителами, называют "антигенами". Глава 7, стр. 386.
ароморфоз - крупное эволюционное преобразование "прогрессивного" характера. По определению И. И. Шмальгаузена, ароморфоз - это "расширение жизненных условий, связанное с усложнением организации и повышением жизнедеятельности". Стр. 309.
археи - одно из трех надцарств живой природы (наряду с бактериями и эукариотами). Вместе с бактериями составляют группу прокариот. Внешне и по образу жизни похожи на бактерий. Отличаются от последних на молекулярном уровне - нуклеотидными последовательностями генов, строением клеточной мембраны, рибосом, важнейших ферментов и др. Часто встречаются в экстремальных местообитаниях (глубоко под землей, в источниках с очень горячей водой). Глава 2, стр. 94.
АТФ - аденозинтрифосфат, главная "энергетическая молекула" живой клетки. Представляет собой нуклеотид аденозин с тремя присоединенными фосфатами. При отделении одного из фосфатов АТФ превращается в АДФ (аденозиндифосфат) и выделяется энергия, которая может быть использована клеткой для тех или иных полезных "работ". Чтобы синтезировать АТФ из АДФ, нужно затратить энергию. Добыча этой энергии, необходимой для синтеза АТФ, составляет одну из основных "жизненных задач", стоящих перед клеткой. Глава 1, стр. 56, 66.
АТФ-синтазы - разнообразные ферменты, синтезирующие АТФ из АДФ и фосфата с использованием того или иного вида энергии - например, энергии химических связей, выделяющейся в ходе какой-либо окислительно-восстановительной реакции, или энергии электрохимического потенциала на клеточной мембране (за счет разной концентрации ионов по обе стороны мембраны образуется разность потенциалов, которую клетка может использовать для синтеза АТФ). Стр. 93.
аэробы - организмы, которым для жизни нужен кислород,
бактерии - одно из трех надцарств живой природы (наряду с археями и эукариотами). Вместе с археями составляют группу прокариот. Глава 2, стр. 94.
белки - биополимеры, молекулы которых представляют собой цепочки из множества последовательно соединенных аминокислот. Химические свойства белка определяются его первичной структурой, то есть последовательностью аминокислот. Эта последовательность закодирована в молекуле ДНК, причем каждая аминокислота кодируется тремя нуклеотидами, составляющими триплет, или кодон. О свойствах белков см. главу 4, стр 205.
бластомеры - клетки эмбриона на ранних стадиях развития. Образуются в результате дробления зиготы (оплодотворенного яйца) у животных. Глава 5, стр. 280-284.
гамета - половая клетка. Например, сперматозоид или яйцеклетка. Гаметы гаплоидны (содержат одинарный набор хромосом). В результате слияния двух половых клеток (оплодотворения) образуется диплоидная клетка - зигота, гаплоидный - содержащий одинарный набор хромосом. В жизненном цикле животных гаплоидная фаза представлена половыми клетками (гаметами - яйцеклетками и сперматозоидами). В результате слияния двух половых клеток (оплодотворения) образуется диплоидная клетка - зигота. Гаплоидная клетка образуется либо в результате редукционного деления (мейоза) диплоидной клетки, либо в результате обычного деления другой гаплоидной клетки.
ген - участок ДНК (или РНК), кодирующий какой-нибудь белок (при помощи генетического кода) или функциональную молекулу РНК. Ген обычно состоит из кодирующих и некодирующих участков. Некодирующие участки могут выполнять регуляторные функции (промоторы, сайты связывания транскрипционных факторов и др). От них зависит, в каких обстоятельствах и с какой интенсивностью будет работать (транскрибироваться) данный ген.
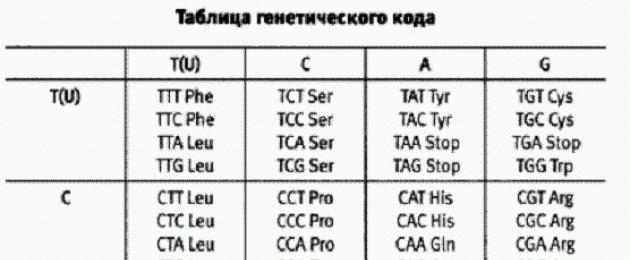
генетический код - универсальный для всех живых существ способ, посредством которого первичная структура белковой молекулы (последовательность аминокислот) "кодируется" в молекуле ДНК (или РНК). Каждая аминокислота кодируется тремя нуклеотида- ми (кодоном, или триплетом). Нуклеотидов в ДНК всего 4, поэтому они могут образовывать 64 разных триплета. Аминокислот в белках всего 20, поэтому генетический код "избыточен": многие аминокислоты кодируются не одним, а несколькими взаимозаменимыми кодонами. Считывание генетической информации проиходит в два этапа: транскрипция и трансляция. Стр. 77.
Стандартный генетический код
Основания ДНК (РНК) A - Adenine A Аденин, Т - Thymine TТимин, (U - Uracil У Урацил), G - Guanine Г Гуанин, С - Cytosine Ц Цитозин
Аминокислоты

геном - в настоящее время под этим термином обычно понимают совокупность всех молекул ДНК, имеющихся в данной клетке и полученных ею "по наследству" от родительской клетки. Изначально термин был предложен для обозначения совокупности всех генов, характерных для данного вида живых существ. Это было в те времена, когда структура ДНК и генетический код еще не были расшифрованы и никто не знал, что в хромосомах помимо собственно "генов" имеется много некодирующих участков. Когда говорят о геноме многоклеточного организма (например, о геноме человека), имеют в виду тот геном, который характерен для половых клеток или оплодотворенного яйца. В соматических клетках многоклеточного организма геном может претерпевать изменения и поэтому быть не совсем таким, как в половых клетках. У РНК-содержащих вирусов геном "сделан" не из ДНК, а из РНК.
генотип - совокупность наследственной информации, полученной организмом от родителей. Или, иначе, совокупность всей наследственной информации, содержащейся в геноме. Обычно генотипом называют только информацию, записанную в форме последовательности нуклеотидов в молекулах ДНК. Некоторая часть наследственной информации передается потомству другими способами (например, с молекулами РНК, с белками цитоплазмы, в виде эпигенетических модификаций ДНК), но все это обычно в понятие "генотип" не включают,
герма - "генеративная" часть организма, передающая свой наследственный материал потомству. У животных это половые клетки и их предшественники. См. также "Сома". Глава 4, сюжет "Появлению многоклеточности мешают обманщики", стр. 225.
гетерозиготный - имеющий два разных аллельных варианта данного гена (см. аллель),
гетеротрофы - организмы, питающиеся готовой органикой, не способные превращать неорганический углерод в органические соединения. Они являются по сути дела нахлебниками автотрофов: они целиком и полностью зависят от производимых ими органических соединений. Глава 2, стр. 95.
гипермутирование соматическое - внесение многочисленных изменений (мутаций) в определенные участки генома соматических клеток. Осуществляется специализированными белками с целью создания методом "проб и ошибок" нового гена с определенными свойствами. Например, таким образом создаются гены новых защитных белков - антител - в ходе выработки приобретенного иммунитета. Глава 7, стр. 388.
гомозиготный - имеющий два одинаковых аллельных варианта данного гена (см. аллель).
горизонтальный обмен генами (горизонтальный перенос) - обмен генетическим материалом между организмами (в отличие от обычной вертикальной передачи генов от родителей потомкам). Широко распространен у прокариот и одноклеточных эукариот (протистов). У многоклеточных эукариот встречается реже, но тоже играет важную роль. Глава 8, раздел "Горизонтальный обмен генами", стр. 420.
градуалистическая эволюция - постепенная, идущая путем отбора небольших изменений. См. раздел "Постепенно или скачками?" в главе 6, стр. 357.
дивергенция - расхождение видов (или признаков) в процессе эволюции. Глава 8, стр. 428.
диплоидный - содержащий двойной набор хромосом. Диплоидная клетка образуется либо в результате деления (митоза) другой диплоидной клетки, либо в результате слияния двух гаплоидных половых клеток (оплодотворение).
ДНК - биополимер, молекула которого представляет собой цепочку из множества последовательно соединенных дезоксирибонуклеотидов (см. врезку "ДНК и РНК - хранители наследственной информации" в главе 1). Обычно две комплементарные цепочки ДНК объединяются вместе, образуя двойную спираль.
ДНК-полимераза - фермент, осуществляющий репликацию (копирование, размножение) молекул ДНК. Стр. 78.
домен - функциональная часть (блок) белковой молекулы. Белковая молекула может содержать один или несколько разных доменов, выполняющих разные функции,
зигота - диплоидная клетка, образовавшаяся из слияния двух гаплоидных половых клеток. Например, оплодотворенное яйцо, значимые и незначимые нуклеотидные замены - как известно, каждая аминокислота в молекуле белка кодируется тремя нуклеотидами в молекуле ДНК. Однако для построения белков используется всего 20 аминокислот, тогда как возможных триплетов - комбинаций из трех нуклеотидов - насчитывается 64. Поэтому говорят, что генетический код "избыточен". В результате большинство аминокислот кодируется не одним, а несколькими разными триплетами. Из-за этого некоторые нуклеотидные замены в кодирующей части гена не приводят к замене аминокислоты в белке (см. генетический код). Такие замены называют незначимыми или синонимичными,
иммуноглобулины - обширная группа (надсемейство) белков, основная функция которых состоит в специфическом распознавании и связывании других молекул. К этой группе относятся, в частности, антитела. Белки надсемейства иммуноглобулинов играют важную роль не только в иммунной системе, но и в межклеточных взаимодействиях, поддержании целостности многоклеточного организма, индивидуальном развитии и т. д. Глава 9, раздел "Взаимное узнавание", стр. 488.
импринтинг геномный - модификация наследственного материала без изменения первичной структуры (последовательности нуклеотидов) ДНК, в частности, путем метилирования нуклеотидов. Может приводить к изменению активности тех или иных генов и, как следствие, к наследственному изменению фенотипа без изменений генотипа. Глава 8, стр. 458.
ингибитор - вещество, останавливающее (замедляющее, подавляющее) какой-либо химический (биохимический, биологический) процесс.
интроны - некодирующие вставки в генах. Обильны в генах эукариот, редки в генах прокариот. Во время транскрипции ген считывается целиком, вместе с интронами. Получившаяся "незрелая" матричная РНК подвергается сплайсингу ("нарезке"), в ходе которого интроны удаляются, стр. 153.
канализированность эволюции - ограниченность числа возможных ("разрешенных") эволюционных преобразований, придающая эволюции частичную предсказуемость. См. сюжет "Пути эволюции предопределены на молекулярном уровне" в главе 4, стр. 213.
кодон - последовательность из трех нуклеотидов, кодирующая одну аминокислоту (см. генетический код),
комплементарность - свойство нуклеотидов, из которых состоят ДНК и РНК, связываться только с определенными (комплементарными) нуклеотидами другой (противоположной) цепочки ДНК или РНК. Нуклеотид А связывается с Т (или У, если речь идет об РНК), Г - с Ц. Свойство комплементарности лежит в основе процессов репликации (копирования) ДНК и РНК, транскрипции, трансляции и др. См. врезку "ДНК и РНК - хранители наследственной информации" в главе 1. Глава 7, стр. 374.
конверсия генов - целенаправленное изменение генов (их нуклеотидных последовательностей), приводящее к превращению одного аллеля в другой. Может осуществляться, например, путем замены участков гена другими, похожими, участками или с помощью перекомбинирования генетических фрагментов-заготовок. Стр. 390.
конъюгация - своеобразный половой процесс у прокариот и инфузорий. Два одноклеточных организма соединяются, обмениваются наследственным материалом и расходятся. Глава 7, раздел "Контролируемая перестройка генома у инфузорий". Стр. 392.
креационизм - вера в божественное сотворение живых существ. Существует очень много версий креационизма, но почти все они сходятся в одном - в отрицании факта эволюции. Существуют версии, претендующие на "научность", однако в действительности креационизм не является научной теорией, в частности из-за отсутствия проверяемых следствий. Подробная и весьма корректная информация о креационизме приведена в статье "креационизм" в русской Википедии: http://ru.wikipedia.org/wiki/ .
мейоз - "редукционное деление" эукариотической клетки, в результате которого число хромосом сокращается вдвое. Из диплоидной родительской клетки (с двойным набором хромосом) получаются четыре гаплоидных клетки с одинарным набором хромосом. У животных таким путем образуются половые клетки - яйцеклетки и сперматозоиды. Стр. 95, 216.
мембрана (клеточная, или плазматическая) - полупроницаемая оболочка, окружающая живую клетку. У всех живых существ основу мембраны составляют два слоя молекул из класса липидов (жиров), но липиды эти могут быть разными. У бактерий и эукариот мембранные липиды представляют собой эфиры глицерина и жирных кислот, а у архей - эфиры глицерина и терпеноидных спиртов. Стр. 67.
метаболизм - обмен веществ. Совокупность всех химических процессов, происходящих в организме,
метагеномный анализ - метод изучения разнообразия организмов (чаще всего - микробов). Берут пробу из какой-либо среды (будь то морская вода или содержимое человеческого кишечника), выделяют оттуда всю ДНК и секвенируют (определяют последовательность нуклеотидов). Затем по этим нуклеотидным последовательностям, используя имеющиеся генетические банки данных, определяют, какие организмы присутствуют в пробе. См. сюжет "Кишечная микрофлора превращает человека в "сверхорганизм" в главе 3, стр. 171.
метаногены - хемоавтотрофные прокариоты (археи), выделяющие в качестве конечного продукта жизнедеятельности метан (СН 4). Глава 2, стр. 101.
метилирование ДНК - прикрепление метильных групп (-СН 3) к некоторым нуклеотидам в молекуле ДНК, осуществляемое специальными ферментами ДНК-метилтрансферазами. Используется, в частности, как один из способов регуляции активности генов. Глава 7, стр. 399. Глава 8, стр. 457.
митоз - деление эукариотической клетки, в результате которого из одной родительской клетки получаются две дочерние с таким же числом хромосом, как и у родительской. Например, митоз диплоидной клетки приводит к формированию двух диплоидных клеток. Стр. 216.
митохондрии - органеллы эукариотической клетки, ответственные за кислородное дыхание. Являются прямыми потомками симбиотических прокариот из группы альфапротеобактерий. Глава 3.
мобильные генетические элементы (МГЭ) - фрагменты ДНК, способные перемещаться с места на место в пределах генома. К ним относятся транспозоны и ретротранспозоны. Глава 8, раздел "Вирусы и мобильные генетические элементы", стр. 438.
модификация (адаптивная) - (адаптивное, то есть приспособительное, полезное) изменение строение клетки или многоклеточного организма, происходящее при неизменном геноме. В основе (адаптивных) модификаций лежит регуляция работы генов: в ответ на те или иные внешние стимулы (сигналы) одни гены начинают работать (экспрессироваться) сильнее, другие слабее. Стр. 241-253.
модификационная изменчивость - изменения строения клетки или многоклеточного организма, основанные не на мутациях, а на модификациях (в том числе адаптивных); разнообразие особей, возникающее на основе таких изменений, морфология - форма, строение, структура организма, а также раздел биологии, изучающий все перечисленное,
мутация - изменение последовательности нуклеотидов в молекуле ДНК (или РНК, если речь идет об РНК-содержащих вирусах). Мутации возникают в силу разных причин - ошибок репликации, перемещений мобильных генетических элементов, жесткого излучения, воздействия химически активных веществ и т. д. Существуют также механизмы "целенаправленного" мутирования, например, при формировании генов новых защитных белков в ходе иммунного ответа. Глава 7, стр. 373.
мутация гомеозисная - наследственное изменение, в результате которого свойства одних частей тела проявляются у других частей. В основе гомеозисных мутаций лежат изменения генов - ключевых регуляторов развития, таких как Нох -гены. Например, у мухи могут вырасти ноги на сегментах брюшка, где в норме их не должно быть, или лишние крылья на заднем сегменте груди. У человека тоже иногда встречаются гомеозисные мутации (например, лишняя пара ушных раковин на шее). У некоторых пород домашних коз такие "лишние уши" даже стали нормой. Стр. 355.
макромугация - крупная мутация, меняющая строение организма настолько, что он может (гипотетически) сразу превратиться в другой вид. См. раздел "Постепенно или скачками" в главе 6. Стр. 357.
мутагенез - появление мутаций.
нуклеотид - составной элемент, мономер, "кирпичик" молекулы РНК (рибонуклеотид) и ДНК (дезоксирибонуклеотид). Каждый нуклеотид состоит из трех частей. Первая из них - фосфорная кислота (фосфат) - неорганическое вещество, которого довольно много в земной коре и океанах. Вторая - сахар рибоза (РНК) или дезоксирибоза (ДНК). Третья - азотистое основание. В состав РНК входит четыре азотистых основания - А (аденин), У (урацил), Г (гуанин) и Ц (цитозин); соответственно, существует четыре вида рибонуклеотидов - аденозин, уридин, гуанозин, цитидин. В состав ДНК тоже входят четыре азотистых основания. Вместо урацила используется тимин (Т); остальные три основания такие же, как в РНК. Глава 1, стр. 56, 75.
онтогенез - индивидуальное развитие организма. Например, развитие многоклеточного животного из оплодотворенного яйца. См. главу 5.
пептиды - обычно так называют белки с короткими молекулами (состоящими из небольшого числа аминокислот),
пластиды - органеллы растительной клетки, отвечающие за фотосинтез. Являются потомками симбиотических цианобактерий. Глава з, стр. 144.
полиморфизм - разнообразие вариантов. Разнообразными могут быть, например, варианты генов (аллели) в популяции - тогда говорят о полиморфном гене; варианты какого-нибудь морфологического признака (такого как окраска) - тогда говорят о полиморфном признаке. Словосочетание "полиморфный вид" означает, что особи, составляющие вид, не одинаковы, разнообразны. Все существующие виды полиморфны, но степень полиморфизма может быть разной,
полиплоидный - содержащий множество хромосомных наборов в одной клетке (см. также гаплоидный и диплоидный),
прокариоты - живые организмы, не имеющие клеточного ядра. Их геном находится прямо во внутренней среде клетки (цитоплазме) и обычно имеет вид единственной кольцевой молекулы ДНК (кольцевой хромосомы). У прокариот нет настоящего полового размножения, точнее говоря, в их жизненном цикле отсутствует фаза образования половых клеток и их попарного слияния в клетку с двойным набором хромосом - зиготу. У прокариот также нет внутриклеточных органелл, окруженных двойными мембранами, - митохондрий и пластид. К прокариотам относятся бактерии и археи. Глава 2, стр. 94.
промотор - участок ДНК, к которому прикрепляется фермент РНК-полимераза перед началом транскрипции. Промотор входит в состав "регуляторной области" гена. Ген без промотора не может быть "считан" (транскрибирован). От свойств промотора зависит, как и когда будет работать ген. Стр. 382.
простейшие - то же, что протисты.
протисты - общее наименование всех одноклеточных эукарот.
псевдогены - неработающие гены, вышедшие из строя в результате мутаций или изначально "молчащие" (например, ретропсевдогены, образовавшиеся в результате деятельности обратной транскриптазы и не имеющие промотора). Стр. 390, 455.
пунктуалистическая эволюция - идущая по принципу "прерывистого равновесия". Длительные периоды относительной стабильности вида чередуются с краткими периодами быстрых изменений. См. раздел "Постепенно или скачками?" в главе 6, стр. 357.
рекомбинация - обмен участками между молекулами ДНК (или РНК). Стр. 157, 377, 387, 405, 432.
репарация - "починка" ДНК, исправление всевозможных ошибок (мутаций, разрывов), возникающих в молекуле ДНК в силу различных причин. Репарация осуществляется специальными ферментами. Глава 9, стр. 502-504.
репликация - удвоение, размножение, копирование. Обычно термин применяется к процессу копирования молекул ДНК. В результате репликации из одной двухцепочечной молекулы ДНК ("двойной спирали") получаются две точно такие же. Иногда в процессе репликации происходят ошибки - мутации. Глава 7, стр. 375.
репродуктивная изоляция - невозможность (или ограниченная возможность) представителей двух разных групп организмов скрещиваться между собой и (или) давать плодовитое потомство. Репродуктивная изоляция считается одним из важных критериев вида. Если представители двух группировок (популяций) не скрещиваются между собой, то это, скорее всего, разные виды. Если они делают это часто и охотно и производят полноценное потомство - значит, они относятся к одному и тому же виду. Впрочем, нельзя абсолютизировать этот критерий.
ретровирусы - группа РНК-содержащих вирусов, которые при помощи механизма обратной транскрипции встраивают копию своего генома в геном хозяйской клетки и размножаются вместе с ним. Родственны ретротранспозонам. Стр. 441-442.
ретропсевдоген - участок ДНК с последовательностью нуклеотидов, идентичной кодирующей части какого-нибудь гена, обычно неактивный, неработающий, который возник в результате деятельности обратной транскриптазы. Глава 8, стр. 455.
ретротранспозон - мобильный генетический элемент, размножающийся при помощи обратной транскрипции. Глава 8, сюжет "Вирусы и мобильные элементы: кто от кого произошел",
рецептор - белок, который избирательно распознает определенное вещество (называемое лигандом), связывается с ним и тем или иным способом "сообщает" о случившемся другим белкам или клеткам. Обычно рецепторные белки располагаются на клеточной мембране, пронизывая ее насквозь. Снаружи от мембраны находится рецепторная часть белковой молекулы. Если она "поймает" лиганд, это приводит к изменению структуры той части молекулы, которая находится по другую сторону мембраны - в цитоплазме. По этому изменению клетка "узнает", что химический сигнал получен. Глава 9, раздел "Взаимное узнавание".
рибозим - молекула РНК с каталитической (ферментативной) функцией, способная осуществлять активную "работу" в клетке. Стр. 58.
рибосома - молекулярная "машинка" для трансляции (синтеза белка на матрице мРНК). Состоит из рРНК и рибосомных белков. Стр. 70.
РНК - рибонуклеиновая кислота, биополимер, молекула которого представляет собой цепочку из множества последовательно соединенных рибонуклеотидов (см. врезку "РНК" в главе 1, стр. 56). матричная (мРНК) - РНК, образующаяся в результате транскрипции (считывания гена). "Незрелая" мРНК содержит интроны и другие некодирующие фрагменты, которые затем удаляются в ходе сплайсинга. Стр. 75, 77.
транспортная (тРНК) - функциональная РНК (рибозим), играющая ключевую роль в "прочтении" генетического кода в ходе трансляции (синтеза белка). Подносит нужные аминокислоты к рибосоме, осуществляющей синтез белка. См.: Антикодон. Стр. 61.
рибосомная (рРНК) - функциональная РНК (рибозим), входящая в состав рибосом. Играет ключевую роль в процессе трансляции (синтеза белка). Стр. 59, 70.
РНК-полимераза - фермент, синтезирующий молекулу РНК на матрице. Матрицей может служить другая молекула РНК или ДНК. В первом случае говорят об РНК-зависимых РНК-полимеразах, во втором - о ДНК-зависимых. Глава 1, раздел "РНК обзаводится помощниками", стр. 70. сайт связывания транскрипционного фактора - короткий участок ДНК, к которому может прикрепиться белок - транскрипционный фактор (ТФ). Сайты связывания ТФ находятся в регуляторных областях генов. Разные ТФ имеют разные сайты связывания. Степень специфичности варьируется: одни ТФ прикрепляются только к строго определенной последовательности нуклеотидов, другие довольствуются расплывчатым "мотивом", в котором только некоторые нуклеотиды являются ключевыми. Глава 8, раздел "Вирусы и мобильные генетические элементы", стр. 438.
секвенирование - экспериментальное определение последовательности нуклеотидов в молекуле ДНК или РНК.
симпатрическое видообразование - разделение исходного вида на два (или более) при обитании на одной и той же территории без физических изолирующих барьеров. См. сюжет "Эволюция на островах идет параллельными путями" в главе 6, стр. 335.
синапс, синаптический контакт - специализированная зона контакта между отростками нервных клеток и другими клетками, нервными, мышечными или иными, обеспечивающая передачу информационного сигнала,
синтетическая теория эволюции (СТЭ) - система эволюционных взглядов, оформившаяся в середине XX века в результате синтеза дарвиновских идей и достижений генетики. Представляет собой важный этап развития эволюционного учения. Некоторые генетические и эволюционные закономерности приобрели в СТЭ характер "нерушимых догм", что потребовало в дальнейшем внесения многочисленных изменений и уточнений в теорию. "Развенчанию" этих догм посвящены многие разделы данной книги. Стр. 16.
сома - "вегетативная" часть организма, которая при размножении обычно не передает свой наследственный материал потомству. Например, у животных "соматическими" являются все клетки, кроме половых. Однако при вегетативном размножении (например, у растений) сома передает свои гены потомству. См. также Герма. Глава 4, сюжет "Появлению многоклеточности мешают обманщики". Стр. 225.
сперматоцит - мужская половая клетка животных в период ее роста и созревания. Стр. 485.
сплайсинг - процесс посттранскрипционной обработки мРНК, в ходе которого происходит удаление интронов. В результате "незрелая" мРНК с интронами превращается в "зрелую" мРНК без интронов, которую уже можно использовать в качестве матрицы для синтеза белка (трансляции). альтернативный сплайсинг - см. одноименную врезку в главе 9, стр. 491.
строматолиты - слоистые минеральные образования, формирующиеся в результате жизнедеятельности микробных сообществ. Стр. 90, 258.
сульфатредукторы - хемоавтотрофные бактерии, живущие за счет реакции восстановления сульфатов. Потребляют сульфаты и восстановитель (например, молекулярный водород), выделяют восстановленные соединения серы (например, сероводород). Стр. 108-116.
теломеры - концевые участки хромосом у эукариот. У прокариот хромосомы не линейные, а кольцевые, поэтому у них нет тело- мер. При каждой репликации теломеры укорачиваются, поэтому на определенных этапах жизненного цикла их приходится восстанавливать - достраивать при помощи специальных ферментов теломераз. Согласно одной из теорий, укорачивание теломер в течение жизни многоклеточного организма является причиной старения (так называемая "теломерная теория старения"). На самом деле, скорее всего, причины старения гораздо сложнее и разнообразнее. Стр. 154.
трансгенный (организм) - генетическая химера, организм, в геном которого были вставлены чужеродные гены. Например, инсулин для диабетиков сегодня производится в промышленных количествах трансгенными бактериями - кишечными палочками, которым пересадили человеческий ген инсулина. Стр. 400.
транскрипция - "считывание генетической информации", синтез РНК на матрице ДНК. Осуществляется ферментом ДНК-зависимой РНК-полимеразой. Полученная молекула РНК в дальнейшем может быть использована для синтеза белка (трансляции). Стр. 77.
обратная транскрипция - синтез ДНК на матрице РНК, переписывание генетической информации из РНК в ДНК. Осуществляется ферментом обратной транскриптазой. Используется некоторыми вирусами (ретровирусами) и ретротранспозонами для встраивания в геном хозяина, а также эукариотами - для восстановления кончиков хромосом (теломер). Путем обратной транскрипции образуются также ретропсевдогены.
транскрипционный фактор - белок, регулирующий экспрессию (активность, уровень транскрипции) каких-либо генов. Транскрипционный фактор узнает определенную последовательность нуклеотидов в регуляторной области гена и прикрепляется к ней. Это в свою очередь либо облегчает, либо, наоборот, затрудняет работу РНК-полимеразы - фермента, осуществляющего транскрипцию (считывание) генов. Глава 8, раздел "Вирусы и мобильные генетические элементы". Стр. 446.
трансляция - синтез белка. Осуществляется особыми молекулярными "машинками" - рибосомами, причем в качестве "матрицы" используется мРНК, полученная в результате транскрипции. Каждые три нуклеотида мРНК (см. кодон) кодируют одну аминокислоту. Аминокислоты присоединяются к синтезируемой молекуле белка по одной при помощи тРНК. Стр. 59.
транспозон - мобильный генетический элемент, перемещение и размножение которого осуществляются при помощи фермента транспозазы без участия обратной транскрипции. Глава 8, стр. 445.
фенотип - строение организма, совокупность всех морфологических, физиологических, биохимических и прочих признаков,
фермент - белок, выполняющий каталитическую функцию, то есть осуществляющий (катализирующий) какую-то химическую реакцию. Молекулы РНК, обладающие такими свойствами, называют рибозимами.
фиксация углерода (фиксация CO 2) - включение неорганического углерода (источником которого является обычно углекислый газ С0 2) в состав органических соединений. См. сюжет "Самая главная химическая реакция" в главе 2, стр. 102.
филогения - родственные отношения между группами живых организмов ("кто от кого произошел") и их изучение,
фотоавтотрофы - автотрофы, получающие необходимую для жизни энергию из солнечного света. Глава 2, стр. 95. аноксигенные фотоавтотрофы в ходе фотосинтеза выделяют не кислород, а другие "отходы жизнедеятельности", чаще всего серу или сульфаты. Нуждаются в восстановленных соединениях (чаще всего в сероводороде). Глава 2, стр. 95. оксигенные фотоавтотрофы в ходе фотосинтеза выделяют кислород. Не нуждаются в восстановленных соединениях серы, в качестве "донора электрона" при фотосинтезе используют обычную воду. Глава 2, стр. 95.
хемоавтотрофы - автотрофы, получающие необходимую для жизни энергию из какой-либо окислительно-восстановительной реакции. Глава 2, стр. 95.
хромосома - молекула ДНК, несущая наследственную информацию, в комплексе с разнообразными структурными белками, которые обеспечивают ее укладку, упаковку и т. п. У прокариот обычно имеется одна кольцевая хромосома, редко две, кроме того, у них часто встречаются дополнительные маленькие кольцевые хромосомы, называемые плазмидами. У эукариот имеется несколько линейных, не замкнутых в кольцо хромосом, цитоплазма - внутренняя среда живой клетки, ограниченная клеточной мембраной.
экзон - кодирующий участок гена. У эукариот большинство генов содержит несколько экзонов, между которыми располагаются интроны. В ходе сплайсинга из незрелой мРНК удаляются интроны, а экзоны "склеиваются" вместе. Глава 9, раздел "Взаимное узнавание", стр. 491.
экосистема - природный комплекс, образованный живыми организмами и средой их обитания, связанный в единое целое обменом веществ и энергии. Например, пруд или лес вместе со всеми своими обитателями,
экспрессия гена - то же, что "работа" гена. Если ген не работает (не подвергается транскрипции), говорят, что он не экспрессируется. Если количество считываемых с гена мРНК увеличивается, говорят об усилении экспрессии, и т. д.
эндемичный - обитающий только в данном месте (районе),
эндогенные ретровирусы - геномы ретровирусов, встроенные в геномы высших организмов и передающиеся по наследству вместе с остальными генами. Глава 8, стр. 454.
эпигенетическое наследование - передача наследственной информации, записанной не в виде последовательности нуклеотидов в молекулах ДНК, а иными способами, например, при помощи метилирования нуклеотидов, метилирования и ацетилирования гистонов - белков, на которые "наматывается" ДНК в клетке. Глава 8, раздел "Эпигенетическое наследование". Стр. 457.
эукариоты - одно из трех надцарств живой природы (наряду с бактериями и археями). Эукариоты - организмы, в чьих клетках есть ядро и окруженные двойной мембраной органеллы: митохондрии, служащие для кислородного дыхания, и пластиды, служащие для фотосинтеза (последние характерны только для растительных клеток). Доказано, что митохондрии и пластиды являются потомками симбиотических бактерий (см. главу 3). К эукариотам относятся разнообразные одноклеточные формы, обычно называемые простейшими или протистами (амебы, жгутиконосцы, инфузории, радиолярии и др.), а также многоклеточные - грибы, растения и животные. В жизненном цикле эукариот есть чередование гаплоидной и диплоидной фаз: пара гаплоидных (с одинарным набором хромосом) половых клеток сливается, образуя диплоидную (с двойным набором хромосом) клетку - зиготу. Это слияние двух половых клеток называют оплодотворением. Затем в какой-то момент происходит редукционное деление, или мейоз, в результате которого из диплоидной клетки образуются четыре гаплоидные. Глава 2, стр. 94.
АЛЕКСАНДР МАРКОВ РОЖДЕНИЕ СЛОЖНОСТИ
В процессе исторического развития одни виды вымирают, другие изменяются и дают начало новым видам. Что же собой представляют виды? Существуют ли виды реально в природе?
Впервые термин "вид" ввел английский ботаник Джон Рей (1628- 1705). Шведский ботаник К. Линней рассматривал вид в качестве основной систематической единицы. Он не был сторонником эволюционных воззрений и считал, что виды со временем не изменяются.
Ж. Б. Ламарк отмечал, что различия между некоторыми видами очень незначительны, и в этом случае выделить виды довольно сложно. Он сделал вывод о том, что виды в природе не существуют, а систематика придумана человеком для удобства. Реально существует только особь. Органический мир представляет собой совокупность особей, связанных между собой родственны ми узами.
Как видно, взгляды Линнея и Ламарка на реальное существование вида были прямо противоположными: Линней считал, чтo виды существуют, они неизменны; Ламарк отрицал реальное существование видов в природе.
В настоящее время общепринята точка зрения Ч. Дарвина: виды реально существуют в природе, но постоянство их относительно; виды возникают, развиваются, а затем либо исчезают, либо изменяются, порождая новые виды.
Вид - это надорганизменная форма существования живой природы. Он представляет собой совокупность морфологически и физиологически сходных особей, свободно между собой скрещивающихся и дающих плодовитое потомство, занимающих определенный ареал и обитающих в сходных экологических условиях. Виды различаются по многим критериям. Критерии, по которым особи относятся к одному виду, представлены в таблице.
Критерии вида
При определении принадлежности особи к какому-либо виду нельзя ограничиваться лишь одним критерием, а необходимо использовать всю совокупность критериев. Так, не возможно ограничиться только морфологическим критерием , поскольку особи одного вида могут различаться внешне. Например, у многих птиц - воробьев, снегирей, фазанов самцы внешне значительно отличаются от самок.
В природе у животных широко распространен альбинизм, при котором в клетках отдельных особей в результате мутации нарушается синтез пигмента. Животные с такими мутациями имеют белую окраску. Глаза у них красные, потому что в радужной оболочке нет пигмента, и сквозь нее просвечивают кровеносные сосуды. Несмотря на внешние отличия, такие особи, например белые вороны, мыши, ежи, тигры, относятся к своим видам, а не выделяются в самостоятельные виды.
В природе существуют внешне почти неразличимые виды-двойники. Так, раньше малярийным комаром называли фактически шесть видов, похожих внешне, но не скрещивающихся между собой и различающихся по другим критериям. Однако из них только один вид питается кровью человека и разносит малярию.
Процессы жизнедеятельности у разных видов часто протекают очень сходно. Это говорит об относительности физиологического критерия . Например, у некоторых видов арктических рыб интенсивность обмена веществ такая же, как и у рыб, обитающих в тропических водах.
Нельзя использовать и один молекулярно-биологический критерий , так как многие макромолекулы (белки и ДНК) обладают не только видовой, но и индивидуальной специфичностью. Поэтому по биохимическим показателям не всегда можно определить, к одному или разным видам относятся особи.
Генетический критерий также не универсален. Во-первых, у разных видов число и даже форма хромосом могут быть одинаковыми. Во-вторых, в одном виде могут быть особи с разным числом хромосом. Так, у одного вида долгоносика имеются диплоидные (2п), триплоидные (Зп), тетраплоидные (4п) формы. В-третьих, иногда особи разных видов могут скрещиваться и давать плодовитое потомство. Известны гибриды волка и собаки, яка и крупного рогатого скота, соболя и куницы. В царстве растений межвидовые гибриды встречаются довольно часто, а иногда бывают и более отдаленные межродовые гибриды.
Нельзя считать универсальным и географический критерий , так как ареалы многих видов в природе совпадают (например,ареал даурской лиственницы и душистого тополя). Кроме того, существуют виды-космополиты, которые распространены повсеместно и не имеют четко ограниченного ареала (некоторые виды сорных растений, комаров, мышей). Ареалы некоторых быстро расселяющихся видов, таких, как домовая муха, изменяются. У многих перелетных птиц различаются ареалы гнездовий и зимовки. Экологический критерий не является универсальным, так как в пределах одного ареала многие виды обитают в очень разных природных условиях. Так, многие растения (например, пырей ползучий, одуванчик) могут жить и в лесу, и на пойменных лугах.
Виды реально существуют в природе. Они относительно постоянны. Виды можно различить по морфологическому, молекулярно-биологическому, генетическому, экологическому, географическому, физиологическому критериям. При определении принадлежности особи к тому или иному виду следует учитывать не один критерий, а весь их комплекс.
Вам известно, что вид состоит из популяций. Популяция представляет собой группу морфологически сходных особей одного вида, свободно скрещивающихся между собой и занимающих определенное место обитания в ареале вида.
Для каждой популяции характерен свой генофонд - совокупность генотипов всех особей популяции. Генофонды разных популяций даже одного вида могут различаться.
Процесс образования новых видов начинается внутри популяции, то есть популяция является элементарной единицей эволюции. Почему же именно популяцию, а не вид или отдельную особь рассматривают как элементарную единицу эволюции?
Особь не может эволюционировать. Она может изменяться, приспосабливаясь к условиям внешней среды. Но эти изменения не эволюционные, так как они не передаются по наследству. Вид, как правило, неоднороден и состоит из ряда популяций. Популяция относительно самостоятельна и может длительное время существовать вне связи с другими популяциями вида. В популяции протекают все эволюционные процессы: у особей возникают мутации, между особями происходит скрещивание, действуют борьба за существование и естественный отбор. В результате генофонд популяции со временем изменяется, и она становится родоначальником нового вида. Именно поэтому элементарная единица эволюции - популяция, а не вид.
Рассмотрим закономерности на следования признаков в популяциях разных типов. Эти закономерности различны для самооплодотворяющихся и раздельнополых организмов. Самооплодотворение особенно часто наблюдается у растений. У самоопыляющихся растений, например гороха, пшеницы, ячменя, овса, популяции состоят из так называемых гомозиготных линий. Чем объясняется их гомозиготность? Дело в том, что при самоопылении увеличивается доля гомозигот в популяции, а доля гетерозигот сокращается.
Чистая линия - это потомки одной особи. Она представляет собой совокупность самоопыляющихся растений.
Начало изучения генетики популяций было положено в 1903 г датским ученым В. Иоганнсеном. Он исследовал популяцию самоопыляемого растения фасоли, легко дающей чистую линию - группу потомков отдельной особи, генотипы которых идентичны.
Иоганнсен взял семена одного сорта фасоли и определил изменчивость одного признака - массы семени. Оказалось, что она варьирует от 150 мг до 750 мг. Ученый высеял отдельно две группы семян: массой от 250 до 350 мг и массой от 550 до 650 мг. Средняя масса семени вновь выросших растений составила в легкой группе 443,4 мг, в тяжелой - 518 мг. Иоганнсен сделал вывод, что исходный сорт фасоли состоит из генетически различных растений.
В течение 6-7 поколений ученый вел отбор семян тяжелых и легких с каждого растения, то есть про водил отбор в чистых линиях. В результате он пришел к выводу, что отбор в чистых линиях не дал сдвига ни в сторону легких, ни в сторону тяжелых семян, значит в чистых линиях отбор не эффективен. А изменчивость массы семян внутри чистой линии является модификационной, ненаследственной и возникает под воздействием условий среды.
Закономерности наследования признаков в популяциях раздельно полых животных и перекрестноопыляемых растений были установлены независимо друг от друга английским математиком Дж Харди и немецким врачом В. Вайнбергом в 1908-1909 гг. Эта закономерность, получившая название закона Харди - Вайнберга, отражает зависимость между частотами аллелей и генотипов в популяциях. Данный за кон объясняет, каким образом в популяции сохраняется генетическое равновесие, то есть число особей с доминантными и рецессивными при знаками остается на определенном уровне.
Согласно этому закону, частоты доминантных и рецессивных аллелей в популяции будут оставаться постоянными из поколения в поколение при наличии определенных условий: высокой численности особей в популяции; свободном их скрещивании; отсутствии отбора и миграции особей; одинаковой численности особей с разными генотипами.
Нарушение хотя бы одного из этих условий ведет к вытеснению одного аллеля (например, А) другим (а). Под действием естественного отбора, популяционных волн и других факторов эволюции особи с доминантным аллелем А будут вытеснять особи с рецессивным аллелем а.
В популяции может измениться соотношение особей с разными генотипами. Предположим, что генетический состав популяции был таким: 20% АА, 50% Аа, 30% аа. Под воздействием факторов эволюции он может оказаться следующим: 40% АА, 50% Аа, 10% аа. Используя закон Харди - Вайнберга, можно вычислить частоту встречаемости любого доминантного и рецессивного гена в популяции, а также любого генотипа.
Популяция - элементарная единица эволюции, так как она обладает относительной самостоятельностью и ее генофонд может изменяться. Закономерности наследования различны в популяциях разных типов. В популяциях самоопыляющихся растений отбор происходит между чистыми линиями. В популяциях раздельнополых животных и перекрестноопыляемых растений закономерности наследования подчиняются закону Харди - Вайнберга.
В соответствии с законом Харди - Вайнберга при относительно постоянных условиях частота аллелей в популяции остается неизменной из поколения в поколение. В этих условиях популяция находится в состоянии генетического равновесия, в ней не происходят эволюционные изменения. Однако в природе нет идеальных условий. Под влиянием факторов эволюции - мутационного процесса, изоляции, естественного отбора и др. - генетическое равновесие в популяции постоянно нарушается, происходит элементарное эволюционное явление - изменение генофонда популяции. Рассмотрим действие различных факторов эволюции.
Один из главных факторов эволюции - мутационный процесс. Мутации были открыты в начале XX в. голландским ботаником и генетиком Де Фризом (1848-1935).
Главной причиной эволюции он считал именно мутации. В то время были известны только крупные мутации, затрагивающие фенотип. Поэтому Де Фриз полагал, что виды возникают в результате крупных мутаций сразу, скачкообразно, без естественного отбора.
Дальнейшие исследования показали, что многие крупные мутации вредны. Поэтому многие ученые считали, что мутации не могут служить материалом для эволюции.
Лишь в 20-х гг. нашего столетия отечественные ученые С. С. Четвериков (1880- 1956) и И. И. Шмальгаузен (1884-1963) показали роль мутаций в эволюции. Было установлено, что любая природная популяция насыщена, как губка, разно образными мутациями. Чаще всего мутации рецессивны, находятся в гетерозиготном состоянии и не проявляются фенотипически. Именно эти мутации и служат генетической ос новой эволюции. При скрещивании гетерозиготных особей эти мутации у потомков могут переходить в гомозиготное состояние. Отбор из поколения в поколение сохраняет особей с полезными мутациями. Полезные мутации сохраняются естественным отбором, вредные - накапливаются в популяции в скрытом виде, создавая резерв изменчивости. Это приводит к изменению генофонда популяции.
Накоплению наследственных различий между популяциями способствует изоляция , благодаря которой между особями разных популяций не происходит скрещивания, а значит, и обмена генетической ин формацией.
В каждой популяции благодаря естественному отбору накапливаются определенные полезные мутации. Через несколько поколений изолированные популяции, обитающие в разных условиях, будут различаться по ряду признаков.
Широко распространена пространственная , или географическая изоляция , когда популяции разделены различными преградами: реками, горами, степями и т. п. Например, даже в близкорасположенных реках обитают разные популяции рыб одного и того же вида.
Различают также экологическую изоляцию , когда особи разных популяций одного вида предпочитают разные места и условия обитания. Так, в Молдавии у желтогорлой лесной мыши образовались лесные и степные популяции. Особи лесных популяций более крупные, пи таются семенами древесных пород, а особи степных популяций - семенами злаков.
Физиологическая изоляция возникает в том случае, когда у особей разных популяций созревание половых клеток происходит в разные сроки. Особи таких популяций не могут скрещиваться. Например, в озере Севан обитают две популяции форели, нерест которых происходит в разные сроки, поэтому они не скрещиваются между собой.
Существует также поведенческая изоляция . Брачное поведение особей разных видов различается. Это препятствует их скрещиванию. Механическая изоляция связана с различиями в строении органов размножения.
Изменение частот аллелей в популяциях может происходить не только под влиянием естественного отбора, но и независимо от него. Частота аллеля может измениться случайным образом. Например, преждевременная гибель особи - единственной обладательницы какого-либо аллеля приведет к исчезновению этого аллеля в популяции. Это явление получило название дрейфа генов .
Важным источником дрейфа генов являются популяционные волны - периодические значительные изменения численности особей популяции. Численность особей изменяется из года в год и зависит от многих факторов: количества пищи, погодных условий, численности хищников, массовых заболеваний и др. Роль популяционных волн в эволюции была установлена С. С. Четвериковым, который показал, что изменение численности особей в популяции влияет на эффективность естественного отбора. Так, при резком сокращении численности популяции могут случайно сохраниться особи с определенным генотипом. Например, в популяции могут сохраниться особи с такими генотипами: 75% Аа, 20% АА, 5% аа. Наиболее многочисленные генотипы, в данном случае Аа, будут определять генный состав популяции до следующей "волны".
Дрейф генов обычно снижает генетическую изменчивость в популяции, главным образом в результате утраты редко встречающихся аллелей. Этот механизм эволюционных изменений особенно эффективен в небольших популяциях. Однако только естественный отбор на основе борьбы за существование способствует сохранению особей с определенным генотипом, соответствующим среде обитания.
Элементарное эволюционное явление - изменение генофонда популяции происходит под влиянием элементарных факторов эволюции - мутационного процесса, изоляции, дрейфа генов, естественного отбора. Однако дрейф генов, изоляция и мутационный процесс не определяют направленности процесса эволюции, то есть выживания особей с определенным, соответствующим среде обитания генотипом. Единственным направляющим фактором эволюции является естественный отбор.
Основные положения эволюционного учения Ч. Дарвина.
- Наследственная изменчивость - основа эволюционного процесса;
- Стремление к размножению и ограниченность средств жизни;
- Борьба за существование - основной фактор эволюции;
- Естественный отбор как результат наследственной изменчивости и борьбы за существование.
ФОРМЫ ЕСТЕСТВЕННОГО ОТБОРА
| ФОРМА ОТБОРА |
ДЕЙСТВИЕ | НАПРАВЛЕННОСТЬ | РЕЗУЛЬТАТ | ПРИМЕРЫ |
| Движущий | При изменении условий существования организмов | В пользу особей, имеющих отклонения от средней нормы | Возникает новая средняя форма, более соответствующая изменившимся условиям | Возникновение у насекомых устойчивости к ядохимикатам; распространение темноокрашенных бабочек березовой пяденицы в условиях потемнения коры берез от постоянного задымления |
| Стабилизи рующий |
В неизменных, постоянных условиях существования | Против особей с возникающими крайними отклонениями от средней нормы выраженности признака | Сохранение и укрепление средней нормы проявления признака | Сохранение у насекомоопыляемых растений размеров и формы цветка (цветки должны соответствовать форме и величине тела насекомогоопылителя, строению его хоботка) |
| Дизруптив ный |
В изменяющихся условиях жизни | В пользу организмов, имеющих крайние отклонения от средней выраженности признака | Образование новых средних норм вместо прежней, переставшей соответствовать условиям жизни | При частых сильных ветрах на океанических островах сохраняются насекомые с хорошо развитыми или с рудиментарными крыльями |
ВИДЫ ЕСТЕСТВЕННОГО ОТБОРА
Задачи и тесты по теме "Тема 14. "Эволюционное учение"."
Проработав эти темы, Вы должны уметь:
- Сформулировать своими словами определения: эволюция, естественный отбор, борьба за существование, адаптация, рудимент, атавизм, идиоадаптация, биологический прогресс и регресс.
- Кратко описать, каким образом та или иная адаптация сохраняется отбором. Какую роль играют в этом гены, генетическая изменчивость, частота генов, естественный отбор.
- Объяснить, почему в результате отбора не образуется популяция идентичных, безупречно адаптированных организмов.
- Сформулировать, что такое генетический дрейф; привести пример ситуации, в которой он играет важную роль, и объяснить, почему его роль особенно велика в небольших популяциях.
- Описать два способа возникновения видов.
- Сравнивать естественный и искусственный отбор.
- Кратко перечислить ароморфозы в эволюции растений и позвоночных, идиоадаптация в эволюции птиц и млекопитающих, покрытосеменных растений.
- Назвать биологические и социальные факторы антропогенеза.
- Сравнивать эффективность потребления растительной и животной пищи.
- Кратко описать черты древнейшего, древнего, ископаемого человека, человека современного типа.
- Указать черты развития и сходства человеческих рас.
Иванова Т.В., Калинова Г.С., Мягкова А.Н. "Общая биология". Москва, "Просвещение", 2000
- Тема 14. "Эволюционное учение." §38, §41-43 стр. 105-108, стр.115-122
- Тема 15. "Приспособленность организмов. Видообразование." §44-48 стр. 123-131
- Тема 16. "Доказательства эволюции. Развитие органического мира." §39-40 стр. 109-115, §49-55 стр. 135-160
- Тема 17. "Происхождение человека." §49-59 стр. 160-172
Эволюционное учение - это наука о причинах, движущих силах, механизмах и общих закономерностях исторического развития живого мира. Эволюцией в биологии называют непрерывное направленное развитие живого мира, сопровождающееся изменением строения и уровней организации разных групп организмов, позволяющее им более эффективно приспосабливаться и существовать в самых различных условиях обитания.
Эволюционное учение является теоретической базой биологии, так как оно объясняет основные особенности, закономерности и пути развития органического мира, позволяет понять причину единства и огромного многообразия органического мира, выяснять исторические связи между разными формами жизни и предвидеть их развитие в будущем. Эволюционное учение обобщает данные многих биологических наук, позволяет понять механизмы и направления изменчивости живой материи и использовать эти знания в практике селекционных работ.
Эволюционное учение возникло не сразу. Оно сложилось как результат длительной борьбы двух принципиально противоположных систем взглядов на жизнь и ее происхождение - идей Божественного сотворения мира и представлений о самозарождении и саморазвитии жизни. На основе этих воззрений в науке сложились два направления - креационизм, развивающий идеи сотворения мира Богом или Высшим разумом, второе - эволюционизм, допускающий возможность самозарождения и саморазвития органического мира. Существовали также представления о вечности жизни в природе.
Уже в древности эти идеи активно обсуждались, и в их развитие внесли большой вклад такие выдающиеся мыслители своего
Додарвиновский период развития эволюционных идей в биологии времени, как Фалес Милетский, Анаксимандр, Анаксимен, Гераклит, Эмпедокл, Демокрит, Платон, Аристотель и многие другие.
В Средние века господствовали в основном идеи креационизма и неизменности мира.
Наиболее крупными учеными додарвиновского периода развития биологии были К. Линней и Ж. Б. Ламарк.
Карл Линней (1707-1778) - выдающийся шведский ученый. Именно он сделал попытку обобщить имевшиеся в то время данные о многообразии органического мира и создать его научную классификацию, изложив свои взгляды по этим вопросам в «Системе природы» (1735). Он является создателем систематики и номенклатуры - наук о принципах классификации и правилах их наименования. Основной таксономической категорией у растений и животных К. Линней считал вид, определяя его как множество сходных особей, воспроизводящих себе подобных. Виды он объединял в роды. В своей системе он выделял пять таксономических категорий разного уровня: класс, отряд, род, вид, разновидность. Для названия видов К. Линней использовал бинарную номенклатуру, то есть двойное наименование - с указанием названий рода и вида (например, мухомор красный, олень благородный и т. п., где первое слово - название рода, а второе - вида). Описания видов и их названия он сделал на латинском языке, принятом тогда в науке. Это намного облегчило взаимное понимание между учеными разных стран, так как в разных языках один и тот же вид может называться совершенно по-разному. Поэтому до сих пор научные названия растений, грибов или любых других организмов принято писать на латинском языке, понятном специалистам разных стран. Всего К. Линней составил описания около десяти тысяч видов растений и животных, объединив их в 30 классов (24 класса растений и 6 классов животных). Однако система К. Линнея была искусственной, основанной на сходстве только внешних признаков. Так, к классу червей он относил кишечнополостных, губок, иглокожих и даже круглоротых, которые сейчас относятся к совершенно разным типам животных. Растения он разделял на классы по наличию или отсутствию цветка, форме цветка и по числу тычинок и пестиков в нем. Но вместе с тем он совершенно правильно отнес человека к отряду приматов. Это было революционным шагом для того времени. Не случайно труд К. Линнея долгое время был запрещен Ватиканом. К. Линней считал виды неизменными, существующими в том состоянии, как их создал Бог. Но он отмечал, что разновидности могут со временем изменяться. Большой заслугой К. Линнея является то, что его систематика фактически отражала результаты эволюции - многообразие организмов от простых форм к более сложным, а таксономические категории впервые определили иерархию и соподчиненность разных групп организмов - от видов до классов.
Очень крупной фигурой в биологии является Жан-Батист Ламарк (1744-1829) - французский ученый, создавший первое целостное эволюционное учение, основы которого он изложил в своем труде «Философия зоологии» (1809). В нем он впервые доказал, что всем видам присуща изменчивость. Основными причинами изменчивости Ж. Б. Ламарк считал влияние внешней среды и стремление живых организмов к совершенству, заложенное в них Богом. Таким образом, по Ламарку, процесс эволюции как бы намечен самим Творцом. Главным механизмом изменчивости видов Ламарк считал упражнение или неупражнение органов. Под влиянием меняющихся условий среды обитания животным приходится менять свои привычки и способы добывания пищи. Например, у жирафа, которому приходится тянуться вверх за листьями деревьев, со временем вытянулась шея (упражнение органа), а у крота, обитающего под землей, произошла потеря зрения (неупражнение органа). Ламарк дал более подробную по сравнению с Линнеем классификацию животных, распределив их по 14 классам. Он отделил позвоночных животных от беспозвоночных. Выделенные им 14 классов животных были разделены по степени усложнения строения на 6 градаций (ступеней усложнения). Так, к 1-ой градации он отнес и полипов, ко 2-ой - лучистых животных и червей, к 3-й - насекомых и паукообразных, к 4-ой - ракообразных, кольчатых червей, усоногих и моллюсков, к 5-ой - рыб и рептилий и к 6-ой - птиц, млекопитающих и человека. Он совершенно справедливо отмечал происхождение высших форм животных от низших и считал, что человек произошел от обезьян. Заслугой Ламарка является также введение в науку терминов «биология» и «биосфера», которые получили впоследствии широкое распространение.
К середине XIX века наука созрела для создания эволюционного учения в биологии. Причин этому было много. Назовем только некоторые из них.
1. Завершение эпохи Великих географических открытий (XV-ХVIII вв.) показало человечеству все многообразие мира.
Ранее, во времена древнего мира, античности, раннего и среднего Средневековья, люди жили в своих городах и селениях, и круг их путешествий ограничивался лишь небольшим набором сопредельных регионов. Это создавало иллюзию об однообразии и стабильности окружающего мира (см. статью: ). Эпоха кругосветных путешествий обнаружила полную несостоятельность этих представлений. Появились многочисленные описания новых земель, их природы и населяющих их племен, растений и животных, которые разрушали привычные воззрения об однородности и неизменности мира.
2. Активная колонизация вновь открытых земель европейцами потребовала составления подробных описаний природы, климата и ресурсов этих районов, что существенно расширяло знания людей о природе . В этой работе принимали участие уже не одиночки-путешественники, а большие массы людей, что способствовало быстрому распространению новых знаний среди широких слоев населения стран Европы.
3. Развитие капитализма в странах Западной Европы ускорило прогресс в технике и научных изысканиях, необходимых для развития промышленности.
4. Интенсивное развитие науки, в свою очередь, ускорило процесс создания эволюционного учения. В это время активно развиваются многие науки о природе, свидетельствующие о ее целостности и определенном развитии: геология, показавшая единство строения минералов и горных пород в разных регионах Земли; палеонтология, накопившая большое количество окаменелостей, давно вымерших растений и животных, что свидетельствовало о древности жизни и смены одних ее форм другими. Кроме того, были обнаружены ископаемые организмы, составляющие явно переходные звенья между ныне существующими и вымершими формами. Эти факты требовали своего объяснения. Успехи сравнительной анатомии выявили общность строения многих групп растений и животных и показывали существование переходных форм между отдельными группами организмов. Цитология выявила общий характер клеточного строения растений и животных. Эмбриология нашла сходство развития зародышей у разных групп животных. Значительные успехи были достигнуты в области селекции растений и животных, свидетельствующие о возможности искусственного изменения их форм и продуктивности.
Все это вместе взятое и подготовило базу и условия разработки эволюционного учения.
Создание эволюционной теории Ч. Дарвина и А. Уоллеса
Основы современной теории эволюции были созданы выдающимся английским ученым-энциклопедистом Чарлзом Дарвином (1809-1882). Независимо от него в это же время работал и пришел к очень близким выводам соотечественник Ч. Дарвина - зоолог Альфред Уоллес (1823-1913).
Научные интересы Ч. Дарвина как натуралиста были чрезвычайно разнообразны: он занимался ботаникой, зоологией, геологией, палеонтологией, теологией, интересовался вопросами селекции и т. п. Большую роль в жизни Ч. Дарвина и формировании его научных идей сыграло кругосветное путешествие в составе экспедиции на корабле «Бигл» в 1831-1836 гг. Там он смог досконально изучить специфику фауны Галапагосских островов, Южной Америки и ряда других районов мира. Уже в этот период у Ч. Дарвина начинают формироваться основные эволюционные идеи и он приближается к открытию принципа дивергенции - расхождения признаков у потомков общего предка как механизма формо- и видообразования. Большую роль в формировании эволюционистских идей Ч. Дарвина сыграло его участие в палеонтологических раскопках в Уругвае, где он познакомился с некоторыми вымершими формами гигантских ленивцев, броненосцев и ряда беспозвоночных. Вернувшись из экспедиции, Ч. Дарвин пишет ряд монографий и выступает с докладами, принесшими ему признание научной общественности и широкую известность.
Анализируя темпы размножения и реальную численность популяций в природе, Ч. Дарвин задался вопросом о причинах вымирания одних форм и выживания других. Для решения этой проблемы он привлекает идеи Томаса Мальтуса (1766-1834) о борьбе за существование в человеческом обществе, изложенные последним в труде «Опыт в законе народонаселения».
Так у Ч. Дарвина родились собственные идеи о роли борьбы за существование в процессах выживания видов в природе и значении естественного отбора как важнейшего фактора, определяющего направление эволюции. Основными механизмами борьбы за существование Ч. Дарвин считал внутри- и межвидовую конкуренцию, а избирательная гибель рассматривалась им как основа естественного отбора. Эти процессы могут ускоряться при пространственной изоляции популяций. Ч. Дарвин совершенно правильно отмечал, что эволюционируют не отдельные особи, а виды и внутривидовые популяции, то есть эволюционный процесс происходит на надорганизменном уровне.
Особую роль в эволюции Ч. Дарвин отводил наследственной изменчивости организмов в популяциях и половому воспроизводству организмов как одному из главных факторов естественного отбора.
Процесс видообразования Ч. Дарвин считал постепенным, он проводил определенные параллели меду естественным и искусственным отбором, приводящим к формированию подвидов, видов и пород или сортов животных и растений. Он подчеркивал также важное значение других наук (палеонтологии, биогеографии, эмбриологии) в доказательствах эволюции. Эти труды были оценены высшей наградой Королевского научного общества. Квинтэссенцией этих сочинений стал труд «Происхождение видов путем естественного отбора или сохранение благоприятствуемых рас (форм, пород) в борьбе за жизнь», изданный Ч. Дарвином в 1859 г. и не потерявший своего значения и в наше время.
Очень похожие взгляды на эволюцию живого мира и ее механизмы представил и А. Уоллес. Даже многие термины в трудах обоих ученых совпали.
А. Уоллес обратился к Ч. Дарвину, как известному эволюционисту, с просьбой просмотреть и прокомментировать его труд. Доклады обоих ученых на эту тему были опубликованы в одном томе Трудов Линнеевского общества, и сам А. Уоллес, и научная общественность единодушно признали приоретет Ч. Дарвина в этих вопросах. Само эволюционное учение долгое время носило имя его основателя - дарвинизм.
Важнейшей заслугой Ч. Дарвина и А. Уоллеса стало то, что они определили главный фактор эволюции - естественный отбор - и тем самым обнаружили причины протекания эволюции живого мира.
Вид как этап эволюционного процесса
Основной эволюционной единицей является вид. Именно вид, по мнению Ч. Дарвина, является центральным звеном эволюционного процесса. Само представление о виде было сформулировано еще в античные времена Аристотелем, который рассматривал вид как совокупность сходных особей. Примерно этих же представлений о виде придерживался и К. Линней, рассматривая его как самостоятельную, дискретную и неизменную биологическую и систематическую структуру. В настоящее время вид рассматривается как реально существующая в природе группа особей. Остальные систематические категории являются в известной мере производными вида, выделяемыми учеными на основании тех или иных признаков (роды, семейства и т. п.).
В современной биологии видом называют совокупность популяций особей, обладающих наследственным сходством морфологических, физиологических и биохимических признаков, свободно скрещивающихся и дающих плодовитое потомство, приспособленных к определенным условиям жизни и занимающих определенную территорию - ареал. Вид - это основная структурная и таксономическая единица в системе живой природы и качественный этап эволюции организмов.
Критерии вида
Каждый вид характеризуется многими признаками, которые носят название критериев вида.
1. Морфологические критерии включают сходство внешнего и внутреннего (анатомического) строения организмов. Морфологические признаки очень изменчивы. Например, деревья, растущие в густом лесу и на открытых пространствах, выглядят по-разному. Иногда в пределах одного вида могут быть особи сильно различающиеся по морфологии. Такое явление носит название полиморфизма. Это может быть связано с наличием разных стадий развития растений и животных, чередованием полового и бесполого поколений и т. п. Так, личиночные и взрослые стадии многих насекомых совершенно не похожи друг на друга. Различаются морфологически стадии медуз и полипов у кишечнополостных, гаметофит и спорофит у папоротникообразных и т. п.
Если особи различаются двумя морфологическими типами, то их называют диморфными (например, половой диморфизм).
Вместе с тем бывают случаи высокого морфологического сходства разных видов. Такие виды носят название видов-двойников.
Не зная всего этого, каждый определенный морфологический тип можно принять за самостоятельный вид или, напротив, разные, но морфологически похожие виды можно неверно отнести к одному виду. Таким образом, морфологический критерий не может быть единственным при определении вида.
2. Генетический критерий вида подразумевает существование вида как целостной генетической системы, составляющей генофонд вида (совокупность генотипов всех особей, относящихся к этому виду).
Каждому виду свойствен определенный набор числа хромосом (у человека, например, диплоидный набор хромосом 2п равен 46), определенная форма, структура, размеры и характер окраски хромосом. У разных видов число хромосом неодинаково, и по этому критерию можно легко различать очень близкие по морфологии виды (виды-двойники). Так были разделены очень похожие друг на друга виды полевок обыкновенных, имеющие 46 и 54 хромосомы, крыс черных (с диплоидными наборами хромосом 38 и 42). Разное число хромосом у разных видов позволяет особям свободно скрещиваться с представителями своего вида, образуя жизнеспособное и плодовитое потомство, но, как правило, оно обеспечивает частичную или полную генетическую изоляцию при скрещиваниях с особями других видов - вызывая гибель гамет, зигот, эмбрионов или же приводя к образованию нежизнеспособного или бесплодного потомства (вспомните, например, мула - бесплодного гибрида осла и лошади, лошака - бесплодного гибрида коня и ослицы).
В настоящее время генетические критерии вида дополнены молекулярными анализами ДНК и РНК (картирование генов, определение последовательности расположения нуклеотидов в молекулах нуклеиновых кислот и т. п.). Это позволяет не только разделять близкие виды, но и определять степень родственной близости или отдаленности разных видов, облегчает проведение филогенетического анализа определенных групп видов, позволяющего выявить родственные связи между разными видами и группами организмов и последовательность их образования.
Однако, несмотря на большие возможности генетических анализов, они также не могут быть абсолютными критериями при определении видов. Например, одинаковые по числу наборы хромосом могут быть у представителей совершенно разных групп растений, грибов или животных. В природе также известны случаи межвидовых скрещиваний с получением жизнеспособных и плодовитых потомков (например, у некоторых видов канареек, зябликов, ив, тополей и др.).
3. Физиологический критерий включает единство всех процессов жизнедеятельности у всех особей одного вида. Это одинаковые способы питания, обмена веществ, размножения и т.п. Это сходство биологических ритмов особей одного вида (периоды активности и отдыха, зимние или летние спячки). Данные признаки также являются важной характеристикой вида, но не единственной.
4. К биохимическим критериям вида можно отнести, например, сходство строения белков, химического состава клеток и тканей, совокупности всех химических процессов, происходящих у всех представителей вида и т.п. К этой же категории признаков можно отнести способность некоторых видов организмов образовывать биологически активные соединения (такие, как антибиотики, токсины, алкалоиды и др.) и любые другие органические вещества (органические кислоты, аминокислоты, спирты, пигменты, углеводы, углеводороды и др.), что широко используется человеком в различных биологических технологиях. Это тоже очень важные признаки вида, дополняющие другие его характеристики.
5. Экологический критерий вида включает характеристику его экологической ниши. Это очень важная характеристика вида, отражающая его место и роль в биоценозах и в биогеохимических круговоротах веществ в природе. Она включает характеристику мест обитания вида, многообразие его биотических связей (место и роль в цепях питания, наличие симбионтов или врагов и т. п.), зависимость от природных факторов (температуры, влажности, освещения, кислотности и солевого состава среды и пр.), периоды и ритмы активности, участие в превращениях определенных или веществ (окисление или восстановление , серы, азота, разложение белков, целлюлозы, лигнина или иных органических соединений и т. п.). То есть экологическая ниша - это полная характеристика того, где вид встречается в природе, когда он активен, в чем и каким образом проявляется его жизнедеятельность. Но и данный критерий не всегда достаточен для определения вида.
6. Географический критерий включает характеристику и величину ареала, занимаемого видом на планете. На этой территории вид встречается и проходит полный цикл развития. Ареал называется первичным, если образование вида произошло именно на этой территории, и вторичным, если территории были заняты видом вследствие случайных миграций, природных катастроф, перемещения человеком и т. п. Ареал может быть сплошным, если вид встречается на всем его пространстве в подходящих местообитаниях. Если ареал распадается на ряд разобщенных и удаленных территорий, между которыми уже невозможны миграции или обмен спорами и семенами, то он называется прерывистым. Выделяют также реликтовые ареалы, занимаемые древними, случайно выжившими видами.
Виды, которые занимают обширные пространства земли и встречаются в разных эколого-географических зонах, называются космополитами, а занимающие лишь небольшие (локальные) территории и не встречающиеся в других местах, получили название эндемиков.
Для видов с обширными ареалами характерна определенная географическая изменчивость, получившая название клинальной изменчивости. У последних видов возможно также существование географических форм и рас и определенных экотипов, приспособленных к конкретным местообитаниям в пределах ареала.
Как уже отмечалось выше, ни один из названных критериев не является достаточным для характеристики видов и последний можно характеризовать только по комплексу признаков.
Популяции
Вид состоит из популяций. Популяцией называется совокупность особей одного вида, обладающих общим генофондом, заселяющих определенную территорию (часть ареала вида) и размножающуюся путем свободного скрещивания. Популяции, в свою очередь, состоят из более мелких групп особей - семей, демов, парцелл и т. п., связанных друг с другом единством занимаемой территории и возможностью свободного скрещивания.
Связь родителей с потомством обеспечивает непрерывность популяции во времени (наличие нескольких поколений особей в популяции), а свободное половое размножение поддерживает генетическое единство популяции в пространстве.
Популяции являются структурной единицей вида и элементарной единицей эволюции.
Популяции - это динамичные группы, они могут объединяться друг с другом, распадаться на дочерние популяции, мигрировать, менять свою численность в зависимости от условий существования, приспосабливаться к определенным условиям жизни, погибать в неблагоприятных условиях.
В пределах ареала вида популяции распределены очень неравномерно. Их будет больше и они будут более многочисленны в благоприятных условиях существования. Напротив, в неблагоприятных условиях и на границах ареала они будут редки и малочисленны. Иногда популяции имеют островной или локальный характер распределения, например, березовые колки на Урале и в Сибири или пойменные рощи и леса в степной зоне.
Число особей, приходящихся на определенную единицу площади или объем среды, носит название плотности популяции. Плотность популяций очень сильно меняется в разные сезоны и годы. Наиболее резко она меняется у мелких организмов (например, у комаров, водорослей, вызывающих цветение водоемов, и т. п.). У крупных организмов численность и плотность популяций более стабильны (например, у древесных растений).
Каждая популяция характеризуется определенной структурой, которая зависит от соотношения в ней особей разного пола (половая структура), возраста (возрастная структура), размеров, разных генотипов (генетическая структура) и т. п. Возрастная структура популяций может быть очень сложной. Наиболее четко это можно наблюдать у древесных растений, где отдельные особи могут существовать многие десятки и даже сотни лет, принимая активное участие в процессах перекрестного опыления. Таким образом, складываются популяции, состоящие из множества родственных друг другу поколений. В других популяциях возрастная структура может быть очень простой, например, у однолетних растений, которые представляют собой одновозрастные группы.
Популяции постоянно изменяются во времени и в пространстве, и именно эти изменения и представляют собой элементарные эволюционные процессы. Вот почему популяции называют элементарной эволюционирующей структурой.
Механизмы и закономерности изменчивости популяций в природе и их генетическую основу подробно изучали крупнейшие российские генетики и эволюционисты А. С. Серебровский (1892-1948) и С. С. Четвериков (1880-1959). Их трудами и работами их последователей созданы основы популяционной генетики.
Основные типы эволюционного процесса
Дивергенция
Дивергенцией Ч. Дарвин называл расхождение признаков в процессе эволюции, приводящее к появлению новых форм или таксонов организмов, происходящих от общего предка. Дивергенция приводит также к преобразованию одних органов тела в другие в связи с выполнением новых функций. Например, после выхода позвоночных животных на сушу их передние конечности претерпели значительные изменения в зависимости от освоения тех или иных типов местообитаний и образа жизни (бегательные у ящериц, волков, кошек, оленей или других, роющие у кротов, крылья у птиц, крылообразные у летучих мышей, хватательные у обезьян, рука у человека, ласты при вторичном освоении водной среды ихтиозаврами, моржами или китообразными и т. д.). Такие органы, имеющие общее происхождение, но выполняющие разные функции, получили название гомологичных. Гомологичными органами являются листья растений, усики гороха, колючки кактусов, шипы барбариса и др.
Конвергенция
Конвергенцией называется независимое возникновение сходных признаков у организмов, имеющих различное происхождение (не родственных друг другу), или у органов, имеющих различное происхождение, но выполняющих сходные функции. Чаще всего конвергенция возникает при заселении сходных типов местообитаний. Например, конвергентное сходство отмечается у крыльев бабочек и рукокрылых, роющих конечностей кротов и медведок, жабр рыб и ракообразных, толчковых ног зайцеобразных и саранчовых и т. п. Но иногда конвергентное сходство возникает под влиянием сходства выполняемых функций, например, удивительная похожесть строения глаз млекопитающих и головоногих моллюсков. Но в любом случае эти органы формируются из разных частей эмбрионов этих животных.
Параллелизм
Параллелизм - это тип эволюции, при котором конвергентное сходство возникает на основе гомологичных органов. Гомологичные органы или морфологические формы, имевшие когда-то общее происхождение, но потом изменившиеся и переставшие быть похожими друг на друга, в новых условиях снова приобретают черты большого сходства. Это вторичное сходство бывших родственных форм. Например, рыбообразная обтекаемая форма вторично возникает при переходе животных от наземного образа жизни к водному. Вспомните похожесть строения акул (первичноводные животные) и ихтиозавров и китообразных (вторично-водные). У кошачьих саблезубость возникала в разное время у разных видов. Причина параллелизма - одинаковое направление естественного отбора и определенная генетическая близость между такими группами организмов.
Филетическая эволюция
Филетическая эволюция, или филогенез, - это такой тип эволюционного процесса, при котором происходит постепенное преобразование одних таксонов в другие без образования боковых ветвей. При этом образуется непрерывный ряд популяций (таксонов), в котором каждый таксон является потомком предыдущего и предком последующего, не имея сестринских таксонов. Этот тип был описан американским исследователем Дж. Симпсоном в 1944 г.
Изучая закономерности эволюции растений, выдающийся российский (советский) генетик Н. И. Вавилов открыл интересные явления, названные им законом гомологических рядов. Этот закон непосредственно вытекает из анализа соотношений и взаимосвязей между разными типами эволюционного процесса и показывает большое сходство эволюционных изменений у родственных групп организмов. Причиной этого является сходство мутаций гомологичных генов в генофондах родственных видов. Поэтому, зная спектр изменчивости одного вида (или рода), можно с большой вероятностью предсказать многообразие форм другого вида (или рода). При этом целые семейства растений могут характеризоваться определенным циклом изменчивости, обнаруживаемой у всех его родов и видов. Так, зная формы изменчивости ячменя, Н. И. Вавилов очень точно предсказал и впоследствии обнаружил сходные формы у пшеницы.
Правила эволюции
Подводя итог изложению процессов микро- и макроэволюции, можно привести несколько общих правил, которым эти процессы подчиняются.
1. Непрерывность и неограниченность эволюции - эволюция возникла с момента образования жизни и будет непрерывно продолжаться, пока существует жизнь.
3. Правило происхождения специализированных групп от неспециализированных. Только неспециализированные, широко приспособленные группы могут дать толчок эволюции и вызвать образование специализированных групп.
4. Правило прогрессирующей специализации групп. Если группа организмов стала на путь специализации, то последняя только углубляется и обратного возврата не происходит (правило Депере).
5. Правило необратимости эволюции. Все эволюционные процессы необратимы, и все новые эволюционные процессы происходят на новой генетической основе (правило Долло). Например, после выхода на сушу ряд животных вернулся к водному образу жизни, сохранив свои эволюционные приобретения. В частности, и ихтиозавры, и китообразные являются вторичноводными животными, но они не превратились в рыб, а остались пресмыкающимися или млекопитающими, сохранив все особенности своих классов.
6. Правило адаптивной радиации. Эволюционное развитие происходит в разных направлениях, способствуя заселению разных сред обитания.
Филогения и систематика как отражение эволюционных процессов
Изучение микро- и макроэволюционных процессов позволяет установить филогенетические (то есть родственные) связи между разными группами живых организмов и определить время появления этих форм.
Филогенезом называют процесс исторического развития группы или конкретного вида. Филогенезом можно также назвать длительный непрерывный ряд множества онтогенезов, отражающий основные эволюционные перестройки. Изучение филогенеза позволяет установить родственные связи между разными таксонами и выяснить механизмы и время эволюционной перестройки определенных групп живых организмов.
Выделяют следующие основные формы филогенеза:
1) монофилия - происхождение разных видов от одного общего предка;
2) парафилия - одновременное образование видов путем синхронной дивергенции предковой формы на два или большее число новых видов;
3) полифилия - происхождение группы видов организмов от разных предков путем гибридизации и/или конвергенции.
Механизмы и способы филогенетических изменений
1. Усиление (интенсификация) функций тела или его органа, например увеличение объема мозга или легких, приведшие к интенсификации их активности.
2. Уменьшение числа функций. Примером может быть преобразование пятипалой конечности у парно- и непарнокопытных животных.
3. Расширение числа функций. Например, у кактусов стебель помимо основных своих функций выполняет функцию запасания .
4. Смена функций. Например, преобразование ходильных конечностей в ласты у вторичноводных млекопитающих (моржей и др.).
5. Замена одного органа другим (субституция). Например, у позвоночных животных хорда заменяется на костный позвоночник.
6. Полимеризация органов и структур (то есть повышение числа однородных структур). Например, эволюция одноклеточных организмов в колониальные и далее в многоклеточные формы.
7. Олигомеризация органов и структур. Это противоположный полимеризации процесс. Например, образование прочного таза путем сращивания нескольких костей.
Систематика как отражение эволюционных процессов
Систематика - наука о положении организмов в общей системе живого мира. Существует множество систем органического мира. Среди них выделяют искусственные системы, учитывающие лишь чисто внешнее сходство между организмами (примером может быть система К. Линнея), и естественные, или филогенетические системы.
Знание систематики необходимо не только с точки зрения определения вида организма (хотя уже это очень важно), но и для понимания его места (а часто и роли) в живом мире, для представления о его происхождении и родственных связях с другими организмами.
Современная систематика основана на тщательном изучении филогенетических связей между разными группами организмов и, по сути дела, во многом отражает основные этапы развития органического мира от простых форм к сложным. Именно так изложен в школьных учебниках материал по систематике растений и животных.
Составной частью систематики является таксономия - наука о принципах классификации живых существ.
Основной таксономической единицей является вид, образующийся в процессе микроэволюции. Родственные виды объединяют в роды, а близкие роды - в семейства. Семейства, имеющие какие-то общие признаки, группируют в порядки (в ботанике) или в отряды (в зоологии). Порядки и отряды объединяют в классы по принципу сходства ряда крупных признаков - одна или две семядоли у цветковых растений, особенности строения и развития у животных (рептилии, птицы, млекопитающие и т. п.).
Сходство некоторых принципиальных признаков позволяет объединять классы в типы (у животных) или отделы (у растений). Пример - цветковые растения (имеют цветок и защищенные плодом семена), хордовые животные (наличие хорды), членистоногие (членистые конечности) и т.п. Причем типы, классы, а часто и порядки могут объединять не только родственные, но и конвергентно сходные формы.
Типы или отделы объединяют в царства по принципу сходства строения и выполняемых функций больших групп организмов. Например, фотосинтезирующие организмы, выделяющие при фотосинтезе кислород, относят к растениям. Царства, как правило, имеют полифилетическое происхождение.
Царства можно объединять в надцарства и империи. В настоящее время выделяют следующие формы жизни.
Неклеточные формы жизни - вирусы.
Клеточные формы жизни:
1) надцарство (или империя) Прокариоты (включает царства Архебактерий и Истинных бактерий); 2) надцарство (или империя) Эукариоты (царства , Животных, Растений и Грибов). Простейших часто объединяют с животными.
Таким образом, крупные систематические категории (царства, типы (отделы), классы, отряды (порядки) являются по сути дела отражением главнейших направлений эволюционного процесса.
Термины эволюционируют помимо и вопреки нашей воле. Поэтому даже самое лучшее определение не может считаться окончательным. В прошлом теории (с оттенком пренебрежения) противопоставляли точному знанию, почерпнутому из Библии или, на худой конец, у Аристотеля. Теория эволюции расценивалась неизмеримо ниже истории сотворения мира, записанной Моисеем со слов бога. Вот почему эволюционисты (например, Дж. Хаксли на праздновании столетия «Происхождения видов») настаивали на том, что эволюция - факт, а не теория (к этому утверждению мы еще вернемся). Сейчас престиж теории вырос настолько, что креационисты склонны считать рассказ Моисея теорией, на равных правах с дарвиновской. Казалось бы, коллизия Моисей-Дарвин помогает понять, где теория, а где мифотворчество. Сам Ч. Дарвин, впрочем, считал, что лишь откорректировал Моисея: бог создал не все, а только немногие исходные виды, предоставив остальное естественному отбору . Некоторые факты, например приспособленность организмов к среде, использовались для подтверждения каждой из трех версий: Моисея (проявление изначальной целесообразности), Дарвина (результат отбора) и Моисея - Дарвина (изначальная целесообразность плюс отбор). Современное науковедение рекомендует не слишком полагаться на подтверждения (которые при желании всегда находятся) Лучше поразмыслить, как можно было бы опровергнуть данную теорию. Если опровержение немыслимо (как, например, опровергнуть идею бога, коль скоро он неисповедим, всемогущ и может организовать даже собственное опровержение?), то теория не принадлежит к числу научных . Ограничив таким образом область науки, мы можем приступить к проверке научных теорий на прочность, пытаясь опровергнуть их с помощью специально спланированных для этой цели экспериментов. В результате наполеоновских войн в руки ученых попали мумии животных древнего Египта, которые можно было сравнить с современными и, таким образом, проверить, была ли эволюция Авторитетная комиссия, в которую входил Ламарк, никакой эволюции не обнаружила. На этом можно бы и поставить точку, если следовать К. Попперу с его опровергающим экспериментом Но может быть, мы просто чего-то недопоняли в проверочном эксперименте? Так, вероятно, думал Ламарк, во всяком случае его веру в эволюционный процесс египетские мумии не поколебали. Другие ученые в большей степени полагались на проверочные эксперименты, считая какую-либо теорию (например наследования приобретенных признаков), в зависимости от результатов, то безоговорочно доказанной, то окончательно опровергнутой. Дело в том, что результаты опыта только кажутся самоочевидными. Они нуждаются в интерпретации, которая в свою очередь неизбежно зависит от предвзятых теоретических представлений. Опровержение столь же теоретически нагружено, как и подтверждение. Даже сама принципиальная возможность опровержения может остаться нераспознанной (или, наоборот, оказаться иллюзорной) и, следовательно, не дает такой четкой демаркационной линии между наукой и ненаукой, как хотелось бы. Эти рассуждения, на первый взгляд, ведут нас к стиранию граней между теорией и мифом, как если бы мифы были просто вчерашними теориями, утратившими убедительность в связи с интеллектуальным взрослением человека, а теории - завтрашними мифами. Но различие все же есть, и заключается оно главным образом в различной способности к развитию (включающему разумеется, элементы опровержения). Моисей апеллировал к авторитету (самому богу), Дарвин- к разуму. Его теория-это логическое решение проблемы приспособленности. В первом случае развитие исключено: миф Моисея нельзя корректировать, не подрывая авторитет первоисточника на котором он зиждется. Во втором оно неизбежно, так как сама теория дает толчок развитию разума и накоплению знаний которое в конечном счете ведет к ее пересмотру. Пора уточнить, что подразумевается под эволюцией. Когда-то эволюцией называли развертывание какой-то программы (в современном понимании-преформизм). Ее противопоставляли: 1) эпигенезу - развитию с новообразованием, 2) революции 3) обратимому развитию, 4) регрессивному развитию (ин- или деволюции). Многие приравнивают эволюцию к образованию видов, но, пожалуй, не стоит что-либо определять через вид - самый противоречивый термин в биологии. Некоторые другие современные значения: нарушение равновесного состояния популяции под действием мутаций и естественного отбора, последовательное изменение признака, историческое развитие группы организмов, или филогенез, развитие жизни на Земле от простейших до высших организмов; но не развитие от оплодотворенной яйцеклетки до взрослого организма, не эпизодическое отклонение от нормы, не изменчивость, повторяющаяся в ряду поколений. О человеке тоже говорят, что его взгляды претерпели определенную эволюцию, имея в виду нечто более длительное, последовательное, серьезное по результатам, чем просто изменение. Очевидно, в современном (и, разумеется, не окончательном) понимании эволюция-это серия последовательных изменений с исторически значимым результатом. Мы не обязаны оговаривать, что изменяется (генотип, признак, популяция, вид), как (непрерывно, прерывисто, скачкообразно, направленно, обратимо - эти эпитеты более или менее условны, как мы еще увидим) и с каким конкретным результатом (видообразование, филогенез, общее развитие жизни и т. п.). Но мы должны признать, что эволюция распознается апостериор-но: изменение, происходящее на наших глязлх, может быть или не быть эволюцией. Вспомнив утверждение, что эволюция - это факт, а не теория (см. выше), заметим, что факт факту рознь (взглянув на термометр, мы говорим о поиышении температуры как о факте, хотя в более строгом смысле слова фактом можно признать лишь расширение ртути; гпяп, его с температурой - скорее теоретическое представление, пошедшее и обиход). Принято думать, что палеонтологические факты подтверждают эволюцию. Однако самыми непримиримыми противниками эволюционизма были в прошлом именно палеонтологи - Ж. Кювье, Л. Агассис, Р. Оуэн и многие другие. Палеонтологическая летопись как таковая - это перечень разрозненных событий, своего рода «хэппенинг». Чтобы составить и:» нее связную историю, нужна руководящая идея. Факты, которыми мы располагаем, заключаются в том,что организмы чрезвычайно разнообразны, приспособлены к тому образу жизни, который они ведут, их жизненное пространство ограничено и они сменяют друг друга в геологической летописи. Объяснения, как мы уже видели, могут быть различными; Теория эволюции состоит в том, что органический мир, каким мы его знаем,- продукт эволюции (в указанном выше смысле). Если же предположить, что мы видим проявления каких-то изначальных свойств живого, не имеющих истории (изначальной целесообразности, например), то такая теория будет неэволюционной или во всяком случае содержащей неэволюционные элементы. Она будет противостоять общей теории эволюции, а не (как часто думают) частной теории естественного отбора, относящейся, как и теории адаптивного изменения под прямым воздействием среды, прогрессивного, необратимого, постепенного, скачкообразного развития и т. д., к числу теорий о теории эволюции, т. е. метатеорий. Это различные решения проблем, возникающих при анализе общей теории эволюции и в свою очередь выдвигающих проблемы для метатеорий следующего яруса.
Александр Болдачев
I. Эволюция и коэволюция
Учитывая, что огромное количество разночтений, недоразумений, противоречий, споров при обсуждении проблем глобального эволюционизма носят сугубо терминологический характер (впрочем, наверное, как и во многих постнеклассических дисциплинах), следующую серию статей я посвящу терминологическим проблемам и мифам, связанным с теорией эволюции.
Глобальный или универсальный?
Термины "глобальный эволюционизм" и "универсальный эволюционизм" используются различными авторами практически как абсолютные синонимы.
Я остановился на слове "глобальный" так как оно наиболее точно отражает представление, что категория "эволюция" относится ко всему Миру как целому, что эволюционный процесс - это и есть форма существования, способ движения Мира. (В этой статье вместо понятия "Вселенная", чтобы избежать излишних физикалистких ассоциаций, я буду использовать слово "Мир"). Термин "универсальный" более подходит для характеристики некоторого принципа (закона), в равной мере применимого к различным обособленным системам.
Это, на первый взгляд, незначительное смысловое различие терминов "универсальный" и "глобальный" отражает противостояние двух эволюционных концепций: (1) коэволюционной и (2) глобально эволюционной (или просто - эволюционной).
Привлечение термина "коэволюция" при рассмотрении глобально-эволюционных процессов, на первый взгляд, кажется вполне естественным и в последнее время стало достаточно популярным.
Значение его предельно просто: коэволюция – это совместная, взаимозависимая эволюция двух или более систем. И тут встаёт вопрос: а возможно ли это? Именно различные ответы на этот вопрос отличают две эволюционные концепции.
Специалисты, развивающие идею коэволюции, естественно вынуждены констатировать, что эволюционирующими можно признать множество относительно независимых систем различного уровня.
Я же придерживаюсь второй точки зрения и считаю, что не существует нескольких эволюционирующих систем и что в каждый момент развития Мира эволюционирующей является только одна, высшая по иерархическому уровню система.
Попробую обосновать свою позицию, основываясь во многом лишь на разрешении элементарных терминологических проблем.
Эволюция и развитие
Несмотря на казалось бы достаточно прозрачное смысловое различие терминов "изменение", "развитие" и "эволюция", существует множество проблем, связанных с неоднозначностью их применения.
О развитии системы, как о чём-то большем, чем о её изменчивости, можно говорить лишь при появлении у системы принципиально новых, ранее не присущих ей определённостей, качеств, степеней свободы и так далее.
Наиболее понятным и наглядным примером развития является рост биологического организма, начиная с единственной клетки до цельного сложноорганизованного существа в зрелом возрасте.
Эволюцией я буду называть процесс развития, при котором возникающие определённости являются новыми, уникальными не только для конкретной системы, но и для всего Мира в целом.
Понятно, что развитие отдельного организма нельзя назвать эволюцией, так как новые для организма структуры, свойства не являются уникальными и уже множество раз повторялись у его предков.
Существенным отличием развития от эволюции является то, что развитие всегда предзадано внешними для самой системы условиями: физическими законами (для космологических и геологических объектов) или, скажем, генетически (для биологических организмов).
Именно некорректное применение термина "эволюция" к космологическим и геофизическим объектам, даёт повод для оппонентов глобально-эволюционного подхода утверждать, что не все эволюционные процессы выстраиваются в единый иерархический ряд и, что не существует единых принципов описания эволюционирующих систем.
Но это чисто терминологическое недоразумение - ни космологические, ни геофизические процессы на данном этапе развития Мира не являются эволюционными. В крайнем случае, можно говорить о развитии космологических объектов (прохождении ими стандартных, определённых существующими законами, стадий), но не об их уникальном эволюционном становлении, то есть появлении у них принципиально новых для Мира определённостей.
Авангард эволюции
Строгая трактовка термина "эволюция", как процесса возникновения принципиально новых для Мира определённостей, позволяет утверждать, что единственной абсолютно эволюционирующей системой является только весь Мир.
Но поскольку движение Мира реализуется в некоторой конкретной локальной (не тождественной Миру) системе, то такую систему также можно назвать эволюционирующей.
В каждый момент времени движения Мира эволюционирующей системой является верхняя, последняя по времени формирования, высшая по уровню развитию ступень в иерархической лестнице.
В настоящее время эволюционирующей является социальная система. Для подчёркивания уникальности, выделенности эволюционирующей системы я ввёл термин "авангардная система" или "авангард эволюции". То есть, можно сказать, что эволюция Мира реализуется возникновением принципиально новых определённостей в процессе функционирования авангардной системы.
Каждая из систем предыдущих иерархических уровней была эволюционирующей, то есть авангардной, в момент своего становления. Конечно, системы ранних уровней участвуют и в последующем эволюционном движении, но лишь как относительно стабильные элементы авангардной системы.
Если в системах предыдущих уровней и происходят принципиальные изменения, то они обязательно связаны с влиянием эволюционирующей авангардной системы.
Так мы можем говорить о выведении новых сортов растений и пород животных, синтезе отсутствующих в природе полимеров, изменениях ландшафта, но все эти принципиально новые для Мира определённости являются следствием функционирования ныне авангардной социальной системы, а не имманентными изменениями биологической, химической и геофизической систем.
Эмпирическим подтверждением существования эволюционных изменений только на высшей (последней) ступени иерархической лестницы является неизменность ранних видов живых организмов - биологическая эволюция осуществлялась только за счёт прироста новых, высших по уровню классов, а не как непрерывная модификация имеющихся видов. Все биологические классы, виды, возникнув в момент, когда они находились на верхней ступени, в дальнейшем уже не претерпевали принципиальных модификаций. Все изменения видов происходили и происходят сейчас либо в рамках изначально определённого уровня, либо вообще в направлении деградации (атрофирование органов, утеря некоторых способностей и так далее).
Миф о коэволюции
Итак, строго определив термины и не найдя подтверждений существования параллельно эволюционирующих систем, я вынужден констатировать, что термин "коэволюция" изначально некорректен. То есть, если под термином "эволюция" понимать лишь появление принципиально новых определённостей, а также, что эволюционное движение свойственно лишь авангардной системе, то использование термина "коэволюция" теряет смысл - буквально, не существует никаких самостоятельно эволюционирующих систем, движение которых мы могли бы рассматривать как коэволюцию.
Можно, конечно, предположить существование множества независимых систем авангардного уровня разнесённых в пространстве, скажем, инопланетных цивилизаций. Однако до факта их контакта невозможно серьёзно обсуждать проблемы коэволюции, а после контакта, мы вынуждены будем рассматривать их как единую систему. Что, по сути, и произошло на нашей планете после окончания эпохи великих географических открытий - ранее независимо развивавшиеся разделённые морями культуры слились в единую цивилизацию.
Иногда говорят о коэволюции, как о согласованном изменении элементов некоторой системы. Однако, в этом случае термин "коэволюция" не означает ничего более, чем просто взаимное согласование элементов. О коэволюции элементов (то есть, об их совместной эволюции) нельзя говорить, даже если они являются частью эволюционирующей системы. Их изменения и в этом случае не эволюционны, а лишь реализуют, отражают эволюционный процесс, оставаясь в рамках своей определённости (смотрите выше замечание о выведении новых пород и синтезе химических элементов).
Особо модным стало использование термина "коэволюция" в социально-экологическом контексте для обозначения стремления к гармоничному взаимодействию цивилизации и биосферы (Моисеев Н. Н.). Такая трактовка термина, конечно, не имеет прямого отношения к парадигме глобального эволюционизма. Серьёзно её можно обсуждать лишь в сферах экологии, социальной этики, политики, просвещения. Коэволюция человека и биосферы с научной позиции (без эмоционально-оценочного экологического пафоса) значит не более, чем просто взаимодействие эволюционирующей социальной системы с окружающей средой. Использование термина "коэволюция" в данном контексте является лишь эффектной заменой таких традиционных понятий из области экологической этики таких, как "гармоничное взаимодействие человека с природой", "соблюдение природоохранных норм" и так далее.
Миф об эволюции биологических видов
А теперь обратимся к изначальному, сугубо биологическому значению термина "коэволюция". Им несколько десятилетий назад стали называть согласованное изменение биологических видов при их сосуществовании в одной экосистеме. По сути, в биологии коэволюция - это не более чем общее название для двух процессов: коадаптации видов и симбиоза (взаимоприспособления видов различных уровней к совместному существованию).
Если для процесса изменения видов использовать традиционный термин "эволюция", тогда коадаптацию и симбиоз вполне корректно можно было бы называть коэволюцией. Но, подходя предельно строго к определению категории "эволюция", мы вынуждены признать, что изменение биологических видов не является эволюционным процессом.
Вид, оставаясь в рамках своей целостности, даже не способен к развитию (обращаю внимание, что я говорю о виде как целом, а не об отдельных организмах). Ведь действительно, нельзя серьёзно считать развитием вида изменение окраски или размеров органов, вызванных приспособлением к окружающей среде. Тем более, что часто (довольно часто) эти изменения носят деградационный характер.
Следовательно, в процессе приспособления вида к среде мы наблюдаем лишь элементарную его изменчивость, движение в рамках заданных определённостей. Но и образование новых видов в рамках одного класса нельзя в полной мере считать развитием и, тем более, эволюционным процессом. Мы же не станем утверждать, что, например, организм тигра более развит по сравнению с организмом камышовой кошки, или слоны являются более эволюционно-прогрессивным видом по сравнению, скажем, с крысами.
Каждый вид млекопитающих (и других классов) идеально приспособлен к своей среде обитания, а морфологические отличия видов в рамках класса не принципиальны с точки зрения уровня развития, и не могут восприниматься как показатели эволюционного изменения.
Этот, самый общий терминологический анализ ставит перед нами вопрос: а является ли биологическая эволюция - эволюцией видов? Терминологически корректнее говорить, что биологическая эволюция реализуется в эволюции классов. И, следовательно, изменчивость видов в ходе естественного отбора нельзя рассматривать, как закон биологической эволюции.
Вот так, незамысловатый анализ эволюционной терминологии не только позволил нам более подробно рассмотреть процесс эволюции Мира, но и по-новому взглянуть на сущность биологической эволюции.
II. Миф об естественом отборе
В развитие начатой темы, эту часть я полностью посвящу логическому анализу современной парадигмы биологической эволюции, истоками восходящей к Чарльзу Дарвину, в частности одному из основополагающих её моментов – принципу естественного отбора (активно применяемого и к другим эволюционирующим системам). И опять же мне придется сделать неутешительный вывод: принцип отбора не только не может рассматриваться как закон эволюции биологического и других иерархических уровней, но и взятый сам по себе является тавтологичным.
Я понимаю, что большинство людей, так или иначе знакомых с теорией Дарвина, жёстко связывают происхождение биологических видов с джентльменским набором: наследственность, изменчивость и отбор. Поверьте, разрушая столь привычное для уха сочетание слов, я преследую лишь единственную цель – попытаться навести порядок в эволюционной терминологии, что, надеюсь, поможет формированию научно состоятельной теории биологической эволюции. А сейчас к делу, то есть к элементарному логическому анализу легендарного принципа отбора!
Недостаточность принципа отбора для объяснения направления эволюции
Начну с предположения (очень разумного), что я ошибаюсь и образование новых видов, как следует из теории Дарвина, происходит исключительно и только вследствие отбора. Для обострения проблемы и большей наглядности рассмотрю сразу процесс образования видов нового класса из видов предыдущего класса. Исходя из того, что теория отбора не содержит каких-либо ограничений на модификацию видов, то есть в ней отсутствуют принципы, запрещающие им постоянно изменяться, логично предположить, что эволюционировать в виды последующего класса должны все или хотя бы некоторое множество видов предыдущего класса. Ну, скажем, из ряда видов пресмыкающихся должен образоваться ряд видов млекопитающих. Если это так, то совершенно непонятно, как в результате случайных изменений видов, приспособленных к обитанию в разных условиях (от пустынь до гор), на разных континентах независимо появились столь одинаковые по функциональному устройству виды последующего класса. Можно предположить два разумных решения этой проблемы: (1) появление видов нового класса от разных видов предыдущего класса не было независимым или (2) все виды следующего класса произошли от одного вида предыдущего. Оба эти варианта вполне могут стать темами для научных исследований. Но для нас важен другой вывод: ни первая, ни вторая из приведённых гипотез, объясняющих функциональную однотипность видов нового класса, не может быть объяснена исходя лишь из принципа отбора. Первый вариант помимо отбора требует существования некоторой не локальной, общей для биосферы закономерности, определяющей направление эволюции. Второй – подразумевает наличие запрета на эволюционное изменение для множества видов класса, что естественно, также не укладывается в рамки концепции отбора.
Адаптивная функция отбора
Вообще-то, для получения вывода, что одного принципа отбора недостаточно для объяснения направленности эволюции, не было необходимости так глубоко копать. Ведь сам по себе, по своей логической сущности отбор не направлен на изменение вида. В условиях стабильной окружающей среды результатом его действия является сохранение стабильности вида. Модификация вида будет происходить только при изменении параметров среды. Из чего следует уже знакомый вывод: отбор нельзя рассматривать как причину, сущность, закон изменения вида – направленность модификации вида определяется изменением внешних условий, а отбор играет лишь вспомогательную адаптивную функцию.
Мутации не задают направление эволюции
Но отбор действует не сам по себе, а в упряжке с двумя другими принципами теории Дарвина: наследственностью и изменчивостью. Понятно, что наследственность выполняет лишь стабилизирующую функцию в жизни вида. А вот изменчивость могла бы претендовать на роль указующего перста эволюции, если бы не признавалась случайной. Следовательно, и в тандеме с изменчивостью отбор не может считаться законом эволюции, определяющим направление модификации видов.
Принцип наименьшего действия как физический аналог принципа отбора
В подтверждение результативности, осмысленности, научной состоятельности принципа отбора часто приводят факт его аналогичности физическому принципу наименьшего действия. Действительно, оба принципа каждый в своей области обеспечивают выбор "лучшей" из множества возможных траекторий движения системы. Но на этом аналогия и заканчивается. Применение принципа наименьшего действия к физической системе возможно только при наличии данных о её состоянии, например, начальных и конечных координат, энергии системы. То есть, сам принцип наименьшего действия не является законом системы, он лишь даёт возможность рассчитать траекторию её движения при известных параметрах системы. А это уже совсем не та роль, которую приписываются принципу отбора, призванному в биологии самостоятельно объяснять направленность и форму реализации эволюционного процесса. По крайней мере, в современной теории биологической эволюции нет никаких других законов, параметров состояния системы и так далее, по отношению к которым отбор мог выступать в качестве принципа поиска оптимального решения, оптимальной траектории движения системы.
Конкуренция в рыночной экономике
Довольно часто можно столкнуться с выдвижением принципа отбора в качестве закона развития элементов социальной системы. Так традиционно, принцип отбора упоминается при описании функционирования рыночной экономики. Но и в роли конкуренции отбор не несёт никакой научной нагрузки, кроме констатации факта приспособления элементов рынка (людей, предприятий) к существующим экономическим условиям. Как и отбор в биологии, конкуренция в экономике не в состоянии объяснить ни появление новых, ни направление модификации старых элементов рынка. Направленность развития экономики, а, следовательно, и её элементов, безусловно, определяется некоторыми общими для всей системы законами, а не банальным принципом выбора из случайных вариантов.
Отбор в сфере научного познания
С таким же успехом принцип отбора применим и в теории научного познания (пример: "Эволюционная эпистемология" К. Поппера, смотрите также мою статью "Тривиальность эволюционной эпистемологии Карла Поппера"). Бесспорно, что и в сфере науки отбор обеспечивает "выживание" лучших теорий. Но точно также он ни в коей мере не объясняет механизм появления новых идей, гипотез. Предположение о том что, теории появляются в результате случайного перебора научных понятий (что вполне логично следует из концепции отбора), нельзя признать состоятельным. Безусловно, что и появление новых теорий, и направление их развития определяются некоторыми (гораздо более содержательными, чем принцип отбора) общими законами функционирования науки и социальной системы в целом.
Какими же должны быть законы эволюционного движения?
Обобщающий вывод можно сделать такой: эволюционная теория может считаться состоятельной, если она будет описывать закономерности развития эволюционирующей системы, как целого. Она должна объяснять возможный спектр новых элементов, новых качеств, а также направленность отбора этих возможностей. Самому же принципу отбора в теории эволюции должно отводиться в лучшем случае место механизма реализации законов, но ни как самого закона.
Тавтологичность принципа отбора
Живучесть теории естественного отбора можно объяснить несколькими причинами: (1) безусловным признанием просвещённой публикой самого факта наличия биологической эволюции и высокой оценкой эвристического значения теории Дарвина, (2) отсутствием какой-либо научной альтернативы (рассматривать не научные, теологические варианты, как не поддающиеся логическому анализу здесь не буду; тех, кого интересует мое мнение на эту тему, отсылаю к статье "Богу - Богово, кесарю - кесарево") и (3) абсолютной тавтологической истинностью принципа отбора.
В качестве закона эволюции принцип отбора имеет научное значение не более чем фраза "получилось так, как получилось". Действительно, на вопросы: "А почему именно этот путь развития имеет место?", "Почему именно такая форма была реализована?" принцип отбора не может ответить ничего более существенного, чем: "они были лучшими", "более приспособленными", "более сильными" и так далее. А самое главное, принцип отбора совершенно не обладает основным свойством научного закона – предсказательной силой. Исходя из отбора, совершенно невозможно прогнозировать развитие системы – можно только констатировать, что "будет так, как будет", или "выживет сильнейший". Ведь если бы биологическая эволюция развивалась совершенно по-другому сценарию, и в результате реализовывались совершенно другие типы организмов, то принцип отбора с той же степенью истинности описал бы и любой другой вариант. Тезис "было, как было, а будет, как будет" опровергнуть нельзя. Но и сделать хоть какой-то научно целесообразный вывод из него тоже невозможно.
Подытоживая рассуждения о месте принципа отбора в эволюционных теориях, ещё раз сформулирую основной пафос моих терминологических изысканий: эволюционное движение системы - это нечто совершенно другое, чем процесс изменчивости и приспособления элементов системы в ходе этого движения. Дарвин "мне друг, но истина дороже".
III. Миф о самоорганизации и саморазвитии
Зададим очень простой вопрос: какой должна быть теория самоорганизующихся систем? Ответ вполне прозрачен: это должна быть теория, способная описать автономное (самостоятельное, не направляемое внешними воздействиями) образование некоторой функционирующей системы из независимых, рассредоточенных элементов. Можно рассмотреть и не такой жёсткий вариант определения, более подходящий для теории саморазвития, чем для теории самоорганизации: это теория, описывающая закономерности появления (формирования, образования) у некоторой автономной системы новых признаков, свойств, функций и так далее, под воздействием только внутренних причин. Казалось бы, вполне корректно сформулированная задача. Вот только есть один вопрос на засыпку: а что же будет описывать эта теория самоорганизации (саморазвития)? Кто мне сможет назвать хоть одну систему (не равную всему Миру), которую можно было бы без натяжек назвать самоорганизующейся или саморазвивающейся?
Первое, что приходит всем в голову – это живые системы. Хорошо, начнём, а вернее продолжим начатый в предыдущих частях разговор о биологической эволюции.
Претендент №1: живой организм
Итак, можно ли считать биологический организм самоорганизующейся, саморазвивающейся системой? Наверное, нет. Появление у отдельного организма новых свойств в период его роста предопределено генетической программой вида. Мы же не будем считать самоорганизацией автомобиля процесс его сборки по чертежам. Хотя, конечно, "сборка" живого организма - это нечто другое. Он сам себя собирает. Но всё же я не стал бы говорить об организме, как о самоорганизующейся системе. Он скорее самовоспроизводится. А это нечто совсем другое. Все его качества предзаданы и определены внешней для него системой - видом. А появление некоторых новых, отличных от общевидовых, качеств носит случайный, мутационный характер, и их нельзя представить как саморазвитие.
Претендент №2: биологический вид
Говорить о саморазвитии вида также не приходится.
Во-первых, мы не можем рассматривать вид как саморазвивающуюся систему, поскольку новые признаки, появляющиеся у вида, отражают, прежде всего, изменения среды, а не некоторое его самостоятельное развитие. Если окружающая среда не изменяется, то и вид остаётся неизменным - действие отбора направлено на поддержание его стабильности.
Во-вторых, поскольку в основе изменений вида лежат случайные мутации (по традиционной трактовке биологической эволюции) или различные внешние воздействия (по альтернативным теориями), то ни о какой внутренней потенции вида к изменениям, которая служила бы движущей силой его саморазвития, говорить не приходиться.
В-третьих, движение биологического вида, как я уже писал ранее, не является развитием в полном смысле этого слова: изменение некоторых морфологических свойств (окраски, размеров и так далее) нельзя рассматривать как приобретение принципиально новых свойств, что только и можно было бы называть развитием. Тем более, что часто приспособление вида к окружающей среде носит деградационный характер.
Признать вид саморазвивающейся системой можно только при условии, что будет точно установлен механизм непосредственного образования вида последующего класса из вида низшего класса (с возникновением новых органов и так далее), да ещё исключительно по внутренним, сугубо видовым причинам (хотя откуда им, этим внутренним причинам, взяться?).
Претендент №3: биосфера (или другие авангардные системы)
Аналогичные рассуждения можно привести и в обоснование "несамостоятельного" развития более крупных биологических таксонов, в частности, классов. Их развитие, безусловно, связано с общими процессами в биосфере - оно отражает и реализует её эволюцию.
Таким образом, мы пришли к вполне естественному выводу, заявленному уже в первой части статьи - эволюционирующими (то есть саморазвивающимися) можно назвать только две системы: Мир и авангардную систему (биосфера была авангардной системой в период биологической эволюции).
Всё сказанное справедливо и для нынешнего авангарда эволюции - социальной системы. Ни один из элементов, ни одну из подсистем социума (людей, предприятия, социальные институты) нельзя рассматривать как саморазвивающиеся, самоорганизующиеся. В своих изменениях они лишь отражают общий процесс эволюции социальной системы.
Даже столь привычное для нас словосочетание "саморазвитие человека" не подразумевает некоторое абсолютное "само-". Человек встроен в социальную систему, и под его "саморазвитием" имеется в виду лишь приобретение внешних человеку знаний, умений без посторонней помощи. Если человека полностью оградить от влияния социума, то его "саморазвитие" ограничится его генетической программой, то есть уровнем животного (хотя, конечно, и высокоразвитого).
Претендент №4 (последний): Мир
Итак, допустим, мы ответили на вопрос: какие системы могла бы описать гипотетическая теория саморазвития (самоорганизации)? Это сам Мир и авангардная эволюционирующая система. Но с первого же взгляда понятно, что они совершенно не подходят под определение самоорганизующихся систем, приведённое в начале текста.
Авангардная система (скажем, биосфера в своё время или социальная система) хоть и приобретает в ходе эволюции новые определённости, но её образование никак не может быть представлено как самоорганизация (самосборка) из некоторых независимых, рассредоточенных элементов. И более того, авангардная система не является абсолютно замкнутой, обособленной от систем предыдущего уровня эволюции. Её развитие обязательно определяется продолжающимся движением (хотя уже не эволюционным) предыдущих систем, которые она включает в себя в качестве элементов. Это - с одной стороны. А с другой - совокупность всех систем более ранних иерархических уровней одновременно является и внешней средой для развития авангардной системы и активно влияет на её формирование. И это жёсткое взаимодействие системы и среды не позволяет рассматривать систему как замкнутую и саморазвивающуюся по своим внутренним причинам. Остаётся сделать вывод, что таковой (замкнутой и саморазвивающейся) системой является только Мир.
САМО- или не САМОразвивается Мир?
Однако тут же встаёт вопрос о правомерности применения терминов "саморазвитие" или "самоорганизация" к такому объекту, как Мир. Определить, является или не является система самоорганизующейся, можно только находясь вне её, то есть с позиции стороннего наблюдателя, в позицию которого мы, естественно, встать не можем. Более того, бессмысленно утверждать, что Мир организуется, поскольку его состояние в каждый конкретный момент времени есть в полной мере организованное. И, конечно, нельзя говорить о некой САМОорганизации, поскольку по определению для Мира никакого "не-САМО" и быть не может.
Конечно, оставаясь в рамках традиционной физики и теории Большого Взрыва, можно представить Мир (Вселенную) как самоорганизующуюся из хаотически распределённых элементов систему. Однако, тем, кто читал мою первую статью о парадигме глобального эволюционизма, понятно, что я могу сказать на эту тему: подобный процесс нельзя считать саморазвитием - он обязательно требует существование внешних Миру физических законов и может быть описан лишь как разворачивание, реализация предзаданного сценария.
А может быть, все же саморазвитие есть на физическом уровне?
Я специально отложил рассмотрение возможности саморазвития на низших иерархических уровнях до более удобного момента, до упоминания проблемы предзаданности. Именно по причине предзаданности сценария изменений все известные нам физические и химические явления и процессы не могут считаться саморазвивающимися. Образование структур или колебательных процессов в открытых нелинейных потоках - не более самоорганизующиеся, саморазвивающиеся явления, чем дифракционная решётка или колебания маятника. Во всех этих случаях мы наблюдаем лишь реализацию жёстко определённого сценария, а не саморазвитие систем или процессов. Более того, во всех нелинейных процессах, представляемых как самоорганизующиеся, образуются не сложные функционирующие системы с множеством взаимодействующих элементов, а лишь регулярные структуры. Описание процесса образования структур - это значительное достижение современной науки, но очень далеко отстоящее от теории саморазвития и самоорганизации сложных систем.
В предыдущем абзаце был брошен камень в огород синергетики, что наверняка не осталось незамеченным просвещённым читателем. Да и весь разговор о самоорганизующихся системах, естественно, подразумевает плавный переход к обсуждению проблем именно этой области современной науки, претендующей на место "теории эволюции и самоорганизации сложных систем" (Е. Н. Князева, С. П. Курдуюмов). Именно этим претензиям синергетики, порождённым, на мой взгляд, элементарной терминологической путаницей, будет посвящена следующая часть статьи о мифах и терминологии эволюционизма. А сейчас вывод по этой части статьи.
Теории самообразования и саморазвития систем не существует и существовать не может по простой причине - отсутствие предмета теории. Для описания систем и процессов, которые я рассматривал в качестве претендентов на роль саморазвивающихся, нужны специальные, довольно не похожие друг на друга теории: теория воссоздания организма по заданной программе, теория изменчивости вида в процессе его адаптации к окружающей среде, теория формирования новых видов в ходе эволюции биосферы, теория образования дипассивных структур в открытых нелинейных средах и так далее, и так далее. Специалисты, работающие над созданием единой (общей) теории самоорганизации и саморазвития сложных систем, наступают на те же грабли, по которым уже десятилетия топчутся физики, работающие над созданием общей теории поля или единой теории физических взаимодействий.
IV. Миф о синергетике как эволюционной теории
Основное, что подтолкнуло меня к написанию данной части статьи – это, на мой взгляд, невзвешенное, некорректное, противоречивое использование терминов «эволюция», «развитие», «саморазвитие», «самоорганизация» в текстах, посвященных молодой, продуктивной науке – синергетике. Я ни в коей мере не хочу принизить ее действительно значимые достижения. Хотелось бы только обратить внимание читателей на необоснованность выводов приверженцев идеи глобальности, новофилософичности синергетической идеологии, делающих смелые экстраполяции результатов этой точной науки в области, далекие от ее сферы приложения.
В двух словах о синергетике
Предметом изучения синергетики (или по другой версии названия - неравновесной термодинамики) являются открытые нелинейные процессы в различных средах (физических, химических, биологических, социальных). Вот, для примера, короткий перечень явлений, которые попадают в сферу интересов синергетики: тепловая конвекция, волны в плазме, погодные и климатические изменения, течение химических и биохимических реакций, колебания численности биологических популяций, демографические процессы и т. п. Нелинейными эти процессы называются потому, что их ход описывается нелинейными уравнениями. Открытость процессов означает, что идут они с постоянным притоком и оттоком энергии, вещества, информации или чего другого, что задействовано в процессе. Неравновесность - что основные эффекты синергетики проявляются в моменты, когда среда или поток находятся в состоянии далеком от равновесия. Неотъемлемая особенность процессов, которые изучает синергетика – это их хаотичность, наличие случайной составляющей, размытость - диссипативность, как говорят синергетики.
Сразу же следует заметить, что синергетика изучает лишь внешние параметры протекания процессов: их скорость, распространение в пространстве и т.д., а не их содержание, способ реализации. То есть специалисты по синергетике не отнимают хлеб от физиков, химиков, экономистов, метеорологов и т.д.
А теперь обратимся к достижениям синергетики, верней к интересным явлениям и эффектам в неравновесных средах, которые она изучает: (1) в открытых нелинейных средах может возникать макроскопическая организованность: волны, регулярные структуры - так называемые диссипативные структуры; (2) для конкретной среды и конкретного процесса возможен дискретный спектр диссипативных структур, что отражает множественность решений нелинейных уравнений; (3) вследствие этой же множественности решений нелинейных уравнений процессы в неравновесной среде при одних и тех же условиях могут протекать по нескольким возможным траекториям (точка, в которой процесс «осуществляет выбор» между возможными траекториями называется точкой бифуркации); (4) образование той или иной структуры, выбор неравновесным процессом той или иной траектории в точке бифуркации зависят от незначительных флюктуаций среды, что позволяет говорить о существенной роли хаоса в протекании неравновесных процессов и формировании структур.
Это, конечно, далеко не весь список. Понятно, что в несколько абзацев не уместить то, чему посвящены тома и тома. Но надеюсь, сказанного достаточно, чтобы понять, о чем идет речь даже впервые прочитавшим слово «синергетика».
А где же эволюция сложных систем?
Для темы нашего разговора, важно то, что в этом кратком изложении основ синергетики мне, без какого либо ущерба для смысла, не понадобилось использовать термины: «сложная система», «развитие», «самоорганизация», «эволюция», которыми постоянно оперируют специалисты по синергетике.
Да и действительно, можно ли потоки или среды (жидкостные, газовые, плазменные, информационные и т.д.) назвать сложными системами? Где элементы, связи, процессы, функционирование?
О каком «развитии» и тем более «эволюции» может идти речь в элементарных процессах горения, теплопроводности, течения химических реакций и т.д.? Где появление принципиально новых качеств, явлений?
Корректно ли назвать «самоорганизацией» появление в среде структур? Тем более, когда структуры не из чего не организуются, а являются лишь регулярными неоднородностями среды, волнами или рядом турбулентностей, и вообще не САМОорганизуются, а формируются потоками.
Конечно, каждая наука свободна в выборе терминологии. Никто не может ограничить использование таких удобных для произнесения фраз: «развитие реакции», «самоорганизация структуры», «многовариантность эволюции процесса». Но именно вольно обиходное применение терминов послужило причиной не всегда обоснованного, на мой взгляд, применения выводов синергетики к сложным системам, в которых реально реализуются процессы развития и эволюции. Именно некорректный перенос терминологии породил такие заключения, что «синергетика – это теория самоорганизации и эволюции сложных систем», «синергетика обосновывает альтернативность путей эволюции» и т.д. (Приведенные фразы взяты не из какого-то конкретного текста, это стандартная риторика, которой предостаточно в статьях, посвященных идеологии синергетики.)
Элементарная проблема редукционизма
Правда, помимо элементарной терминологической неряшливости, некоторым оправданием перескока с разговора о потоках и средах, к обсуждению развития сложных развивающихся систем может служить наличие в последних нелинейных диссипативных процессов. Эти процессы (например, экономические, демографические процессы в социальной системе) возникают в результате хаотичного взаимодействия огромного числа элементов сложной системы и действительно могут быть описаны нелинейными уравнениями в рамках синергетического подхода. Но согласитесь, что формальный расчет некоторых частных процессов в системе никак не означает описание функционирования самой сложной системы. То есть я хочу сказать, что предметом синергетики является не сложная система как таковая, а лишь некоторые процессы в этой системе. Перед нами элементарная проблема редукционизма. К примеру, мы же не будем утверждать, что химия в состоянии описать функционирование биологического организма, только на основании того, что в живом организме протекает множество химических реакций.
Многовариантность эволюции или просто разнообразие форм?
Наличие нескольких решений у нелинейных уравнений означает не более того, что нелинейные процессы в той или иной системе могут иметь несколько вариантов осуществления. Но из этого нельзя сделать вывод, что сама система имеет множество путей эволюции.
Во-первых, априорное наличие нескольких решений ни в коей мере не говорит об эволюции, а является лишь констатацией возможности нескольких предписанных состояний. Эволюцией можно назвать появление нового нелинейного процесса, описываемого новым уравнением с принципиально новыми степенями свободы, а не факт существования у известного уравнения нескольких решений.
Во-вторых, само наличие нескольких решений или вообще какое-либо разнообразие элементов, процессов и т.д. не есть разнообразие эволюционных путей. Опять же, большую достоверность имеет обратное утверждение: единая эволюционная определенность реализуется в многообразии форм. Так, появление нового эволюционного класса животных (например, млекопитающих) реализуется в огромном многообразии видов. Тут, конечно, очень существенную роль играет фиксация терминологии. Если в качестве эволюционного процесса признать появление видов – то эволюция действительно имеет множество путей, а если относить к эволюционным изменениям лишь появление нового класса (что реально является принципиально новой определенностью, новым вариантом организации живого организма, см. предыдущую статью), то следует признать, что в эволюции Мира на каждом этапе реализуется лишь один вариант, но во множестве форм. (Однако не следует принимать это утверждение, как приписывание Миру строгого детерминизма. Я говорю лишь о единичности результата, а не о предписанности конкретной формы его реализации).
В-третьих, мы в принципе не можем определить, является ли эволюция Мира детерминированной или нет по одной простой причине: Мир для нас – это уникальный объект, мы являемся его элементами, и любые наши заверения о вариантах его движения как целого принципиально не проверяемы. Более того, Мир вообще не может быть предметом синергетики, так как он не является открытой средой (потоком).
Хотя, конечно, вывод синергетики о вариантности течения нелинейных процессов и случайности выбора одного из вариантов в точке неустойчивости вполне применим к неуникальным элементам эволюционирующей системы. То есть в случае, если элементы эволюционирующей системы могут реализовываться в нескольких дискретных формах, то процесс выбора одной из них одним конкретным элементом (например, направление развития и форма отдельного экономического предприятия) вполне вписывается в схему синергетического подхода.
Новая философия или все же очередная точка зрения?
Следует четко разделять синергетику, как точную науку, описывающую процессы в открытых нелинейных средах и потоках, и синергетическую естественнонаучную парадигму, переносящую, экстраполирующую точные научные выводы на процесс развития сложных систем, чаще всего на социум и его элементы. Схема переноса вполне привычная. Так, несколько веков назад классическая механика заставила всех говорить о детерминизме и возможности рассчитать все и вся. Теория Дарвина побудила повсюду видеть действие принципа отбора. Теория относительности вызвала рассуждения на тему относительности всяких и в том числе социальных представлений. Квантовая механика инициировала споры о дуализме общественных процессов, кибернетика - об их информационной сущности. Ну а синергетика, решив задачу возникновения структур в неравновесных средах, и наблюдая множество нелинейных процессов в сложных систем, решила, что вот сейчас-то она опишет саморазвитие последних, а там уж рукой подать до формулы (нелинейной, конечно) эволюции Мира.
Каждая новая естественнонаучная парадигма, прилагаемая к любой сложной системе (например, социальной) с какой-то новой стороны освещает ее сущность. Но при этом, следует отдавать себе отчет в том, что ни одна из перечисленных и ни какая другая наука в отдельности не может описать движение эволюционирующей системы как целого. Так, для анализа стационарных связей и механизма саморегуляции необходимо привлечь теорию систем и кибернетику. Если система находится в движении и в ней присутствует потоки случайно распределенных элементов, то для описания этого процесса (и только этого процесса, а не всей системы) возможно использовать синергетику.
Синергетика, практически так же, как и принцип отбора отражает лишь механизм реализации эволюционных процессов, и, безусловно, не может поведать нам об их причинах и направленности, и, конечно же, не может претендовать на роль общей теорию эволюционирующих систем.