Fettsyraoxidation kan vara patologiskt ökad eller patologiskt minskad.
Öka hastigheten av fettsyraoxidation, särskilt med brist på kolhydrater, inträffar:
1. När du äter mat rik på fett.
2. Under fasta.
3. För diabetes.
I detta fall bildas ett stort antal ketonkroppar av acetyl-CoA, som bildas vid β-oxidation av fettsyror i levern. Ansamlingen av ketonkroppar leder till acidos och kallas ketos.
Nedgång hastigheten för fettsyraoxidation observeras vid:
1. Brist på karnitin. Det observeras hos nyfödda, oftare för tidigt födda barn. Det orsakas antingen av en kränkning av biosyntesen av karnitin eller av dess "läckage" i njurarna.
Symtom:
· attacker av hypoglykemi som uppstår på grund av en minskning av glukoneogenesen som ett resultat av avbrott i oxidationen av fettsyror;
· minskning av syntesen av ketonkroppar, åtföljd av en ökning av innehållet av fria fettsyror i blodplasman;
Myasthenia gravis (muskelsvaghet);
· ansamling av lipider.
Behandling: tar karnitin oralt.
2. Minskad aktivitet av karnitinpalmitoyltransferas.
I levern leder det till hypoglykemi och en minskning av innehållet av ketonkroppar i blodplasman.
I muskler - till störning av oxidationen av fettsyror, vilket resulterar i muskelsvaghet och utveckling av myoglobinuri.
3. Dikarboxylsyrauri.
Huvudsymtomet är utsöndringen av C 6 -C 10 dikarboxylsyror och hypoglykemi utvecklas, inte associerat med en ökning av ketonkroppar.
Etiologi: frånvaron i mitokondrierna av acetyl-CoA-dehydrogenas av medelkedjiga fettsyror, som är förkortade till dikarboxylsyror med medelkedjiga kedja, utsöndrade från kroppen.
Förekommer hos människor efter att ha ätit omogna ackee-frukter, som innehåller toxinet hypoglycin, vilket inaktiverar acyl-CoA-dehydrogenas, vilket resulterar i hämning av β-oxidationsprocessen.
5. Zellwegers syndrom (cerebrohepatorenalt syndrom).
Det är en sällsynt ärftlig sjukdom där peroxisomer saknas i alla vävnader. Hos patienter som lider av Zellwegers syndrom ansamlas C26-C28-polyensyror i hjärnan, eftersom på grund av frånvaron av peroxisomer genomgår de inte oxidation av långkedjiga fettsyror.
6. Refsums sjukdomar.
Sällsynt neurologisk sjukdom. Förknippas med en medfödd störning i α-oxidationssystemet, vilket leder till ackumulering av fytansyra i vävnader, vilket blockerar β-oxidationssystemet.
Bestämning av nivån av totala lipider i blodplasma (serum) med hjälp av en färgreaktion med ett sulfofoshovanilinreagens
Totala lipider är ett generaliserat koncept som inkluderar icke-förestrade fettsyror, triglycerider, fosfolipider, fritt och förestrat kolesterol och sfingomyeliner.
Metodens princip: nedbrytningsprodukterna av omättade lipider bildar med reagenset (bestående av svavelsyra, ortofosforsyror och vanillin) en förening, vars färgintensitet är proportionell mot innehållet av totala lipider i blodserumet.
Reagenser:
1. Koncentrerad svavelsyra;
2. Fosforovanilinblandning. 4 volymer koncentrerad ortofosforsyra blandas med en volym 6 g/l vanillinlösning. Blandningen förvaras i en mörk glasbehållare vid rumstemperatur.
3. Triolein standardlösning, 8 g/l.
Framsteg av beslutsamhet
Till 0,02 ml blodserum tillsätt 1,5 ml koncentrerad svavelsyra. Innehållet blandas och placeras i ett kokande vattenbad i 15 minuter. Efter kylning av hydrolysatet, mät upp 0,1 ml (kontrollprov 0,1 ml koncentrerad svavelsyra), som överförs till andra provrör innehållande 1,5 ml fosfovanillinreagens. Efter blandning inkuberas proverna i 50 minuter på en mörk plats vid rumstemperatur. Den optiska densiteten för provet (A 1) och referenslösningen (A 2) mäts på en fotokolorimeter vid en våglängd på 510-540 nm i en kyvett med en skikttjocklek på 10 mm mot kontrolllösningen. Beräkningen görs med hjälp av formeln: .
Normalhalt i blodserum: 4 - 8 g/l.
Klinisk och diagnostisk betydelse. Förändringar i blodnivåerna av de kvantitativa och kvalitativa komponenterna i denna indikator observeras i många sjukdomar och patologiska tillstånd som inte diskuteras i denna handbok. I förhållande till muskelaktivitet observeras en ökning av denna indikator efter långvarig fysisk aktivitet, vilket visar i vilken grad lipidmetabolism ingår i energitillförseln av muskelaktivitet. Dessutom går värdet på denna indikator vanligtvis inte utöver referensgränserna. Mer informativt är att bestämma dynamiken i skift under fysisk aktivitet, komponenterna i denna indikator.
BIOSYNTES AV LIPIDER
Lipidbiosyntes (lipogenes) är nödvändig för att skapa lagringsformer. Lipidbiosyntesen börjar med biosyntesen av fettsyror.
Biosyntes av fettsyror
Fettsyrasyntessystemet är lokaliserat i den lösliga cytoplasmatiska fraktionen av många organ och vävnader, såsom lever, njurar, bröstkörtel och fettvävnad.
Biosyntesen av fettsyror sker med deltagande av:
1. NADPH∙H+;
5. Acetyl-CoA som substrat och palmitinsyra som slutprodukt.
Funktioner av fettsyrabiosyntes
Fettsyrasyntes är inte en enkel vändning av β-oxidationsreaktioner. De viktigaste funktionerna är följande:
1. Syntesen av fettsyror sker i cytoplasman, till skillnad från den nedbrytning som sker i mitokondrierna.
2. Mellanprodukter från fettsyrasyntes är kovalent kopplade till sulfhydrylgrupperna i acyltransferproteinet (ATP).
3. Många enzymer för syntes av fettsyror i högre organismer och människor är organiserade i ett multienzymkomplex som kallas fettsyrasyntetas.
4. Acetyl-CoA i sig används endast som primer.
5. Den växande fettsyrakedjan förlängs genom direkt tillsats av tvåkolkomponenter härledda från acetyl-CoA. Den aktiverade donatorn av tvåkolkomponenter vid förlängningsstadiet är malonyl-CoA. Förlängningsreaktionen utlöses av frigörandet av CO 2 .
6. Rollen som ett reduktionsmedel i syntesen av fettsyror spelas av NADPH·H +.
7. Fettsyrasyntes är en cyklisk process som sker på ytan av fettsyrasyntetas.
8. Förlängning under verkan av fettsyrasyntetaskomplexet upphör vid bildningsstadiet av palmitat (C 16). Ytterligare förlängning och införande av dubbelbindningar utförs av andra enzymsystem.
Stadier av fettsyrabiosyntes
Steg I - transport av acetyl-CoA från mitokondrier till cytoplasman
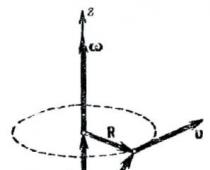
Fettsyror syntetiseras i cytoplasman och acetyl-CoA bildas från pyruvat i mitokondrierna. Det mitokondriella membranet är inte permeabelt för acetyl-CoA, så transporten av acetyl-CoA över membranet säkerställs av speciella mekanismer. Karnitinets roll i transporten av acetyl-CoA är inte stor, eftersom det endast transporterar långkedjiga fettsyror. Detta problem löses genom att syntetisera citrat.
Mitokondrier Cytoplasma
Acetyl-CoA + oxaloacetat acetyl-CoA + oxaloacetat + ADP + Pn
HO - C - COOH-citrat + ATP + HSKoA
CH2 - COOH
Ris. 20. Schema för acetyl-CoA-transport genom mitokondriella membranet
Citrat bildas i mitokondriematrisen genom kondensation av acetyl-CoA och oxaloacetat. Sedan diffunderar det in i cytoplasman, där det klyvs av citratlyas. Således överförs acetyl-CoA och oxaloacetat från mitokondrier till cytoplasman med hjälp av en enda molekyl av ATP.
Källor till NADPH H+ för fettsyrabiosyntes
Oxaloacetat som bildas som ett resultat av överföringen av acetyl-CoA till cytoplasman måste återföras till mitokondrien. Denna process är associerad med genereringen av NADPH·H+. Reaktionen sker i cytoplasman och sker i två steg:
1. Oxaloacetat + NADH + Malat + NAD +
MDH (dekarboxylerande)
2. Malat + NADP + Pyruvat + CO2 + NADPH H+
Det resulterande pyruvatet diffunderar lätt in i mitokondrierna, där det karboxyleras till oxaloacetat av pyruvatkarboxylas (med utgifter för ATP-energi).
Pyruvat + HCO3 - + ATP Oxaloacetat + ADP + Phn
Normal fettoxidation i kroppen är nära relaterad till Krebs-cykeln. Huvudvägen för oxaloacetatbildning är karboxylering av PVK. För att bränna 1,5 g fettsyror krävs 1 g kolhydrater. Därför finns det ett talesätt bland biokemister att "fetter brinner i kolhydraternas lågor."
Oxaloacetatet som syntetiseras i denna reaktion reagerar sedan med acetyl-CoA för att bilda citrat, som oxideras i TCA-cykeln.
Således, för varje molekyl av acetyl-CoA som passerar från mitokondrier till cytoplasman, bildas en molekyl av NADPH·H +. Följaktligen, under övergången av 8 molekyler acetyl-CoA som är nödvändiga för syntesen av palmitinsyra, bildas 8 molekyler av NADPH·H+. Ytterligare 6 molekyler som krävs för denna process genereras i pentosfosfatvägen.
Steg II - bildning av malonyl-CoA.
Det är den första reaktionen i biosyntesen av fettsyror. Katalyseras av enzymet acetyl-CoA-karboxylas. Koenzymet är biotin. Reaktionen består av karboxylering av acetyl-CoA, källan till CO2 är bikarbonat.
C = O + HCO 3 - + ATP E– biotin CH 2 + ADP+H 3 PO 4
acetyl - CoA malonyl - CoA
Ris. 21. Karboxylering av acetyl-CoA (coenzym av acetyl-CoA-karboxylas är biotin)
Malonyl-CoA är i huvudsak aktiverad acetyl-CoA. Energi lagras i förväg i form av en karboxylgrupp och frigörs vid dekarboxylering direkt under biosyntesen av fettsyror. I den vidare biosyntesen av fettsyror används acetyl-CoA som frö, och själva syntesen utgår från malonyl-CoA.
Steg III - biosyntes av fettsyror.
förekommer i levern, njurarna, skelett- och hjärtmusklerna och fettvävnaden. I hjärnvävnad är graden av fettsyraoxidation mycket låg; Den huvudsakliga energikällan i hjärnvävnaden är glukos.
oxidation av fettsyramolekylen i kroppsvävnader sker i β-position. Som ett resultat avskiljs tvåkolsfragment sekventiellt från fettsyramolekylen på sidan av karboxylgruppen.
Fettsyror, som ingår i de naturliga fetterna hos djur och växter, har ett jämnt antal kolatomer. Varje sådan syra från vilken ett par kolatomer elimineras passerar så småningom genom smörsyrasteget. Efter ytterligare en β-oxidation blir smörsyra till acetoättiksyra. Den senare hydrolyseras sedan till två molekyler ättiksyra.
Leveransen av fettsyror till platsen för deras oxidation - till mitokondrierna - sker på ett komplext sätt: med deltagande av albumin transporteras fettsyran in i cellen; med deltagande av speciella proteiner (fettsyrabindande proteiner, FABP) - transport inom cytosolen; med deltagande av karnitin - transport av fettsyror från cytosolen till mitokondrierna.
Processen för fettsyraoxidation består av följande huvudsteg.
Aktiveringfettsyror. Fri fettsyra, oavsett längden på kolvätekedjan, är metaboliskt inert och kan inte genomgå några biokemiska omvandlingar, inklusive oxidation, förrän den är aktiverad. Aktivering av fettsyran sker på den yttre ytan av mitokondriella membranet med deltagande av ATP, koenzym A (HS-KoA) och Mg 2+ joner. Reaktionen katalyseras av enzymet acyl-CoA-syntetas:
Som ett resultat av reaktionen bildas acyl-CoA, som är den aktiva formen av fettsyran.
Man tror att aktiveringen av fettsyra sker i 2 steg. Först reagerar fettsyran med ATP för att bilda acyladenylat, som är en ester av fettsyran och AMP. Därefter verkar sulfhydrylgruppen i CoA på acyladenylatet som är tätt bundet till enzymet för att bilda acyl-CoA och AMP.
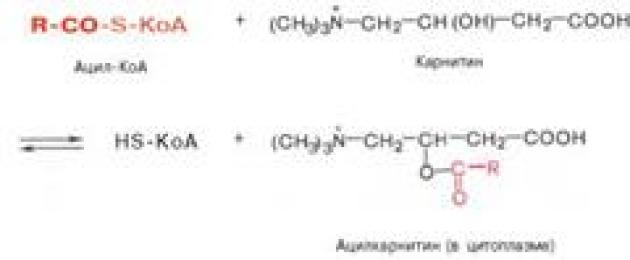
Transportfettsyrorinuti mitokondrier. Fettsyrans koenzymform, precis som fria fettsyror, har inte förmågan att tränga in i mitokondrierna, där deras oxidation faktiskt sker. Karnitin fungerar som en bärare av aktiverade långkedjiga fettsyror över det inre mitokondriella membranet. Acylgruppen överförs från svavelatomen i CoA till hydroxylgruppen av karnitin för att bilda acylkarnitin, som diffunderar över det inre mitokondriella membranet:

Reaktionen sker med deltagande av ett specifikt cytoplasmatiskt enzym, karnitinacyltransferas. Redan på den sida av membranet som är vänd mot matrisen överförs acylgruppen tillbaka till CoA, vilket är termodynamiskt gynnsamt, eftersom O-acylbindningen i karnitin har en hög gruppöverföringspotential. Med andra ord, efter att acylkarnitin passerar genom mitokondriella membranet, inträffar en omvänd reaktion - klyvningen av acylkarnitin med deltagande av HS-CoA och mitokondriellt karnitin-acyltransferas:

Intramitokondriellfettsyraoxidation. Processen för fettsyraoxidation i cellmitokondrier inkluderar flera på varandra följande enzymatiska reaktioner.
Första steget av dehydrering. Acyl-CoA i mitokondrier genomgår först enzymatisk dehydrering, och acyl-CoA förlorar 2 väteatomer i α- och β-positionerna och förvandlas till CoA-estern av en omättad syra. Således är den första reaktionen i varje cykel av acyl-CoA-nedbrytning dess oxidation av acyl-CoA-dehydrogenas, vilket leder till bildandet av enoyl-CoA med en dubbelbindning mellan C-2 och C-3:

Det finns flera FAD-innehållande acyl-CoA-dehydrogenaser, som var och en har specificitet för acyl-CoA med en viss kolkedjelängd.
Skedehydrering. Omättad acyl-CoA (enoyl-CoA), med deltagande av enzymet enoyl-CoA-hydratas, fäster en vattenmolekyl. Som ett resultat bildas β-hydroxyacyl-CoA (eller 3-hydroxyacyl-CoA):

Observera att hydratiseringen av enoyl-CoA är stereospecifik, som hydratiseringen av fumarat och akonitat (se sid. 348). Som ett resultat av hydratisering av trans-A2-dubbelbindningen bildas endast L-isomeren av 3-hydroxiacyl-CoA.
Andra fasendehydrering. Den resulterande p-hydroxiacyl-CoA (3-hydroxiacyl-CoA) dehydreras sedan. Denna reaktion katalyseras av NAD+-beroende dehydrogenaser:

Tiolasreaktion. Under de tidigare reaktionerna oxiderades metylengruppen vid C-3 till en oxogrupp. Tiolasreaktionen är klyvningen av 3-oxoacyl-CoA med användning av tiolgruppen i den andra CoA-molekylen. Som ett resultat bildas en acyl-CoA förkortad med två kolatomer och ett tvåkolsfragment i form av acetyl-CoA. Denna reaktion katalyseras av acetyl-CoA acyltransferas (β-ketotiolas):

Den resulterande acetyl-CoA genomgår oxidation i trikarboxylsyracykeln, och acyl-CoA, förkortad med två kolatomer, går återigen upprepade gånger genom hela β-oxidationsvägen tills bildandet av butyryl-CoA (4-kolförening), som i tur oxideras upp till 2 acetyl-CoA-molekyler
Under en cykel av β-oxidation bildas 1 molekyl acetyl-CoA, vars oxidation i citratcykeln säkerställer syntesen 12 mol ATP. Dessutom bildar det 1 mol FADH 2 och 1 mol NADH+H, under vars oxidation i respirationskedjan det syntetiseras, respektive 2 och 3 mol ATP (5 totalt).
Sålunda, under oxidationen av till exempel palmitinsyra (C16), 7 β-oxidationscykler, vilket resulterar i bildning av 8 mol acetyl-CoA, 7 mol FADH 2 och 7 mol NADH+H. Därför är ATP-utgången 35 molekyler som ett resultat av β-oxidation och 96 ATP som härrör från citratcykeln, vilket motsvarar totalen 131 ATP-molekyler.
FETTSYRA- alifatiska karboxylsyror, av vilka många finns i animaliska och vegetabiliska fetter; i kroppen hos djur och växter fyller fria fettsyror och fettsyror som ingår i lipider en extremt viktig funktion - energisk och plastisk. Omättade fettsyror deltar i människo- och djurkroppen i biosyntesen av en speciell grupp biologiskt aktiva ämnen - prostaglandiner (se). Innehållet av fria och esterbundna fettsyror i blodserum fungerar som ett ytterligare diagnostiskt test för ett antal sjukdomar. Flytande föreningar används i stor utsträckning för framställning av olika tvålar, vid tillverkning av gummi- och gummiprodukter, lacker, emaljer och torkande oljor.
Beroende på antalet karboxylgrupper i molekylen särskiljs en-, två- och flerbasiska flytande föreningar, och beroende på graden av mättnad av kolväteradikalen särskiljs mättade (mättade) och omättade (omättade) flytande föreningar. Baserat på antalet kolatomer i den flytande syrakedjan De är indelade i lägre (C1-C3), mellan (C4-C9) och högre (C10-C26) - Mättade fettsyror har en allmän molekylformel C n H 2 n O 2. Den allmänna formeln för omättade fettsyror beror på antalet dubbel- eller trippelbindningar de innehåller.
Rationell och systematisk nomenklatur används för att beteckna bostäder; Dessutom har många bostadskomplex historiskt etablerade namn. Enligt rationell nomenklatur anses alla flytande föreningar vara derivat av ättiksyra, där väteatomen i metylgruppen i molekylen är ersatt av en kolväteradikal. Enligt den systematiska nomenklaturen kommer namnet på den flytande blandningen från namnet på kolvätet, vars molekyl är uppbyggd av samma antal kolatomer, inklusive kolet i karboxylgruppen, som den flytande syramolekylen (till exempel , propan - propansyra, etan - etansyra, hexan - hexansyra, etc.). Namnet på omättade flytande föreningar anger antalet dubbelbindningar (mono-, di-, tri-, etc.) och lägger till ändelsen "ene". Numreringen av flytande kolatomer börjar med kolet i karboxylgruppen (COOH-) och indikeras med arabiska siffror. C-atomen närmast COOH-gruppen betecknas alfa, den bredvid betecknas beta, och den terminala kolatomen i kolväteradikalen betecknas omega. Dubbelbindningen i en flytande syramolekyl betecknas med symbolen Δ eller helt enkelt ges numret på kolatomen på vilken dubbelbindningen är belägen, vilket indikerar cis- eller trans-konfigurationen av kedjan. Några av de vanligaste bostadskomplexen och deras triviala, rationella och systematiska namn ges i tabell 1.
Fysikaliska egenskaper
Lägre fettsyror är flyktiga vätskor med en stickande lukt, medelstora fettsyror är oljor med en obehaglig härsken lukt och högre fettsyror är fasta kristallina ämnen som är praktiskt taget luktfria.
Endast myrsyra (se), ättiksyra (se) och propionsyra blandas med vatten i alla avseenden; i högre medlemmar av vätskesyraserien minskar lösligheten snabbt och blir slutligen lika med noll. J.-föreningar är mycket lösliga i alkohol och eter.
Smältpunkterna i den homologa serien av flytande kristaller ökar, men ojämnt. Flytande kristaller med ett jämnt antal C-atomer smälter vid en högre temperatur än följande flytande kristaller, som har en C-atom till (tabell 2). I båda dessa serier (med ett jämnt och udda antal C-atomer) minskar skillnaden i smälttemperaturerna för två på varandra följande medlemmar gradvis.
Denna märkliga skillnad mellan flytande föreningar med ett jämnt och udda antal C-atomer i molekylen manifesteras inte bara i smältpunkterna, utan till viss del i de kemiska egenskaperna. och även i deras biol, egenskaper. Således sönderfaller syror med ett jämnt antal C-atomer, enligt G. Embden, vid blödning i levern till aceton, men syror med ett udda antal C-atomer sönderfaller inte.
Flytande kristaller är starkt associerade och även vid temperaturer som överstiger deras kokpunkt visar de två gånger mol. vikt än deras formel antyder. Denna association förklaras av förekomsten av vätebindningar mellan enskilda vätskemolekyler.
Kemiska egenskaper
De kemiska egenskaperna hos flytande föreningar bestäms av egenskaperna hos deras COOH-grupper och kolväteradikaler. I COOH-gruppen är O-H-bindningen försvagad på grund av en förskjutning i elektrontätheten i den dubbla C=O-bindningen mot syre, och därför kan protonen lätt avlägsnas. Detta leder till uppkomsten av en stabil anjon:
Elektronaffiniteten för karbonylresten kan delvis tillfredsställas av den närliggande metylengruppen; väteatomerna är de mest aktiva jämfört med de andra. Dissociationskonstanten för COOH-gruppen av flytande föreningar är 10 -4 -10 -5 M, dvs dess värde är mycket lägre än för oorganiska föreningar. Den starkaste av syrorna är myrsyra. COOH-gruppen av flytande syra har förmågan att reagera i vattenlösningar med alkaliska jordartsmetaller. Salter av högre flytande föreningar med dessa metaller kallas tvålar (se). Tvål har egenskaperna hos ytaktiva ämnen - tvättmedel (se). Natriumtvålar är fasta, kaliumtvålar är flytande. Hydroxyl COOH-grupper av flytande syra kan lätt ersättas med halogen för att bilda syrahalider, som används i stor utsträckning i organiska synteser. När en halogen ersätts med en rest av en annan syra bildas flytande syraanhydrider; när en rest ersätts med en alkohol bildas deras estrar, med ammoniak - amider och med hydrazin - hydrazider. De vanligaste i naturen är estrar av den tribasiska alkoholen glycerol och högre fettsyror - fetter (se). Vätet i alfa-kolatomen i flytande kristaller kan lätt ersättas med halogen för att bilda halogenhaltiga flytande föreningar.Omättade flytande föreningar kan existera i form av cis- och trans-isomerer. De flesta naturliga omättade fettsyror har en cis-konfiguration (se Isomerism). Graden av flytande omättnad bestäms genom jodometrisk titrering av dubbelbindningar. Processen att omvandla omättade fettsyror till mättade kallas hydrering, den omvända processen är dehydrering (se hydrering).
Naturliga fettsyror erhålls genom hydrolys av fetter (deras förtvålning) följt av fraktionerad destillation eller kromatografisk separation av de frigjorda fettsyrorna Icke-naturliga fettsyror erhålls genom oxidation av kolväten; reaktionen fortskrider genom bildningsstadiet av hydroperoxider och ketoner.
Fettsyraoxidation
Som energimaterial används flytande syror i processen för betaoxidation. 1904 lade F. Knoop fram en hypotes som förklarade mekanismen för fettsyraoxidation i djurkroppen.
Denna hypotes byggdes på grundval av fastställandet av de slutliga metaboliska produkterna som utsöndras i urinen efter administrering av ko-fenylsubstituerade fettsyror till djur.I experimenten av F. Knoop, administrering av fenylsubstituerade fettsyror innehållande en Jämnt antal C-atomer till djur åtföljdes alltid av frisättning av fenylättiksyra i urinen, och de som innehöll ett udda antal C-atomer - frisättning av bensoesyra. Baserat på dessa data föreslog F. Knoop att oxidationen av den flytande syramolekylen sker genom att sekventiellt skära av tvåkolsfragment från den från karboxylgruppen (schema 1):
F. Knoops hypotes, kallad teorin om betaoxidation, är grunden för moderna idéer om mekanismen för oxidation av fettsyror Följande metoder och upptäckter spelade en viktig roll i utvecklingen av dessa idéer: 1) införandet av en radioaktiv markör (14 C) in i molekylen av fettsyror för att studera deras utbyte; 2) Munoz och L. F. Leloirs fastställande av det faktum att oxidation av fettsyror genom cellulära homogenat kräver samma kofaktorer som oxidation av pyruvat (oorganiskt fosfat, Mg 2+-joner, cytokrom c, ATP och vilket substrat av Trikarboxylsyracykel - succinat, fumarat, etc.); 3) fastställa det faktum att oxidationen av fettsyror, såväl som substraten för trikarboxylsyracykeln (se Trikarboxylsyracykeln), endast sker i cellens mitokondrier [Lehninger (A.L. Lehninger) och Kennedy (E.P. Kennedy)] ; 4) fastställande av karnitins roll i transporten av fettsyror från cytoplasman till mitokondrier; 5) upptäckt av koenzym A av F. Lipmann och F. Linen; 6) isolering från djurvävnader i renad form av ett multienzymkomplex som ansvarar för oxidationen av fett.
Processen för oxidation av järnsyra består i allmänhet av följande steg.
Fri fettsyra, oavsett längden på kolvätekedjan, är metaboliskt inert och kan inte genomgå några omvandlingar, inklusive oxidation, förrän den är aktiverad.
Aktivering av fettsyror sker i cellens cytoplasma, med deltagande av ATP, reducerade CoA (KoA-SH) och Mg 2+ joner.
Reaktionen katalyseras av enzymet tiokinas:
Som ett resultat av denna reaktion bildas acyl-CoA, som är den aktiva formen av fettsyror.Flera tiokinaser har isolerats och studerats. En av dem katalyserar aktiveringen av fettsyror med en kolvätekedjelängd från C2 till C3, den andra från C4 till C12, och den tredje från C10 till C22.
Transport till mitokondrier. Koenzymformen av fettsyror, liksom fria fettsyror, har inte förmågan att tränga in i mitokondrier, där deras oxidation faktiskt sker.
Det har fastställts att överföringen av den aktiva formen av fettsyror till mitokondrier utförs med deltagande av den kvävehaltiga basen karnitin. Genom att kombinera med fettsyror med hjälp av enzymet acylkarnitintransferas, bildar karnitin acylkarnitin, som har förmågan att tränga in i mitokondriella membranet.
I fallet med palmitinsyra, till exempel, representeras bildningen av palmityl-karnitin enligt följande:
Inuti mitokondriella membranet, med deltagande av CoA och mitokondriellt palmityl-karnitintransferas, inträffar en omvänd reaktion - klyvningen av palmityl-karnitin; i detta fall återvänder karnitin till cellens cytoplasma och den aktiva formen av palmitinsyra, palmityl-CoA, passerar in i mitokondrierna.
Första oxidationssteget. Inuti mitokondrierna, med deltagande av fettsyradehydrogenaser (FAD-innehållande enzymer), börjar oxidation av den aktiva formen av fettsyror i enlighet med teorin om betaoxidation.
I det här fallet förlorar acyl-CoA två väteatomer i alfa- och betapositionerna, och förvandlas till omättad acyl-CoA:
Hydrering. Omättad acyl-CoA fäster en vattenmolekyl med deltagande av enzymet enoylhydratas, vilket resulterar i bildandet av beta-hydroxyacyl-CoA:
Det andra steget av fettsyraoxidation, liksom det första, fortskrider genom dehydrogenering, men i detta fall katalyseras reaktionen av NAD-innehållande dehydrogenaser. Oxidation sker på platsen för beta-kolatomen med bildandet av en ketogrupp vid denna position:

Det sista steget av en komplett oxidationscykel är klyvningen av beta-ketoacyl-CoA genom tiolys (och inte hydrolys, som F. Knoop antog). Reaktionen sker med deltagande av CoA och enzymet tiolas. En acyl-CoA förkortad med två kolatomer bildas och en molekyl ättiksyra frigörs i form av acetyl-CoA:

Acetyl-CoA genomgår oxidation i trikarboxylsyracykeln till CO 2 och H 2 O, och acyl-CoA går återigen igenom hela vägen för beta-oxidation, och detta fortsätter tills nedbrytningen av acyl-CoA, som alltmer förkortas med två kolatomer kommer att leda till bildandet av den sista acetyl-CoA-partikeln (schema 2).
Under betaoxidation, till exempel palmitinsyra, upprepas 7 oxidationscykler. Därför kan det övergripande resultatet av dess oxidation representeras av formeln:
C15H31COOH + ATP + 8KoA-SH + 7NAD + 7FAD + 7H2O -> 8CH3CO-SKoA + AMP + 7NAD-H2 + 7FAD-H2 + pyrofosfat
Den efterföljande oxidationen av 7 molekyler NAD-H 2 ger bildandet av 21 molekyler ATP, oxidationen av 7 molekyler FAD-H 2 - 14 molekyler av ATP och oxidationen av 8 molekyler acetyl-CoA i trikarboxylsyracykeln - 96 molekyler ATP. Med hänsyn till en molekyl ATP som användes i början på aktiveringen av palmitinsyra, kommer det totala energiutbytet för den fullständiga oxidationen av en molekyl palmitinsyra i en djurorganism att vara 130 ATP-molekyler (med fullständig oxidation av en glukos). molekyl, endast 38 ATP-molekyler bildas). Eftersom förändringen i fri energi under fullständig förbränning av en molekyl palmitinsyra är 2338 kcal, och den energirika fosfatbindningen av ATP kännetecknas av ett värde på 8 kcal, är det lätt att beräkna att cirka 48% av den totala potentialen energin av palmitinsyra under dess oxidation i kroppen används för att återsyntetisera ATP, och resten går tydligen förlorad som värme.
En liten mängd fettsyror i kroppen genomgår omega-oxidation (oxidation vid platsen för metylgruppen) och alfa-oxidation (vid platsen för den andra C-atomen). I det första fallet bildas en dikarboxylsyra, i det andra - en fettsyra förkortad med en kolatom Båda typerna av oxidation sker i cellens mikrosomer.
Fettsyrasyntes
Eftersom vilken som helst av oxidationsreaktionerna av fettsyror i sig är reversibla, har det föreslagits att biosyntesen av fettsyror är en omvänd process till deras oxidation. Detta trodde man fram till 1958, tills det fastställdes att i duvleverextrakt kunde syntesen av fettsyror från acetat endast ske i närvaro av ATP och bikarbonat. Bikarbonat visade sig vara en absolut nödvändig komponent, även om det i sig inte ingick i fettsyramolekylen.
Tack vare forskningen från S. F. Wakil, F. Linen och R. V. Vagelos på 60-70-talet. 1900-talet Det visade sig att den faktiska enheten för fettsyrabiosyntes inte är acetyl-CoA, utan malonyl-CoA. Den senare bildas genom karboxylering av acetyl-CoA:

Det var för karboxyleringen av acetyl-CoA som bikarbonat-, ATP- och Mg2+-joner krävdes. Enzymet som katalyserar denna reaktion, acetyl-CoA-karboxylas, innehåller biotin som en protesgrupp (se). Avidin, en biotinhämmare, hämmar denna reaktion, såväl som syntesen av fettsyror i allmänhet.
Den totala syntesen av fettsyror, till exempel palmitinsyra, med deltagande av malonyl-CoA kan representeras av följande ekvation:
Som följer av denna ekvation kräver bildningen av en molekyl palmitinsyra 7 molekyler malonyl-CoA och endast en molekyl acetyl-CoA.
Processen för fettsyntes har studerats i detalj i E. coli och några andra mikroorganismer. Det enzymsystem som kallas fettsyrasyntetas i E. coli består av 7 individuella enzymer associerade med den sk. acylöverföringsprotein (APP). AP B isolerades i sin rena form och dess primära struktur studerades. Mol. vikten av detta protein är 9750. Det innehåller fosforylerat pantetein med en fri SH-grupp. AP B har inte enzymatisk aktivitet. Dess funktion är endast förknippad med överföringen av acylradikaler. Reaktionssekvensen för syntes av fettsyror i E. coli kan presenteras enligt följande:
Därefter upprepas reaktionscykeln, beta-ketocapronyl-S-ACP med deltagande av NADP-H 2 reduceras till beta-hydroxikapronyl-S-ACP, den senare genomgår dehydrering för att bilda omättad hexenyl-S-ACP, som sedan reducerad till mättad kapronyl-S-ACP, med en kolkedja två atomer längre än butyryl-S-APB, etc.
Således är sekvensen och karaktären av reaktioner i syntesen av fettsyror, som börjar med bildningen av beta-ketoacyl-S-ACP och slutar med fullbordandet av en cykel av kedjeförlängning med två C-atomer, omvända reaktioner av oxidation av fettsyror, men syntesvägarna och oxidationen av vätskor korsar sig inte ens delvis.
Det var inte möjligt att detektera ACP i djurvävnader. Ett multienzymkomplex som innehåller alla enzymer som behövs för syntesen av fettsyror har isolerats från levern.Enzymerna i detta komplex är så hårt bundna till varandra att alla försök att isolera dem individuellt har misslyckats. Komplexet innehåller två fria SH-grupper, varav en, som i ACP, tillhör fosforylerat pantetein, den andra till cystein. Alla reaktioner av syntesen av fettsyror sker på ytan eller inuti detta multienzymkomplex. Fria SH-grupper i komplexet (och möjligen hydroxylgruppen i serinet som ingår i dess sammansättning) deltar i bindningen av acetyl-CoA och malonyl-CoA, och i alla efterföljande reaktioner spelar pantetein-SH-gruppen i komplexet samma roll eftersom SH-gruppen ACP, dvs. deltar i bindningen och överföringen av acylradikalen:
Det fortsatta reaktionsförloppet i djurorganismen är exakt detsamma som presenterats ovan för E. coli.
Fram till mitten av 1900-talet. man trodde att levern är det enda organ där syntesen av fettsyror sker.Då fann man att syntesen av fettsyror även sker i tarmväggen, i lungvävnaden, i fettvävnaden, i benmärgen, i l aktiverande bröstkörtel, och även i kärlväggen. När det gäller cellulär lokalisering av syntes finns det anledning att tro att den sker i cellens cytoplasma. Det är karakteristiskt att hl syntetiseras i levercellers cytoplasma. arr. palmitinsyra. När det gäller andra fettsyror är det huvudsakliga sättet för deras bildning i levern att förlänga kedjan baserat på redan syntetiserad palmitinsyra eller fettsyror av exogent ursprung, mottagna från tarmarna. På detta sätt bildas till exempel flytande föreningar innehållande 18, 20 och 22 C-atomer. Bildandet av fettsyror genom kedjeförlängning sker i cellens mitokondrier och mikrosomer.
Biosyntesen av fettsyror i djurvävnader är reglerad. Det har länge varit känt att levern hos svältande djur och djur med diabetes långsamt införlivar 14C-acetat i magen. Samma sak observerades hos djur som injicerades med överskott av fett. Det är karakteristiskt att i leverhomogenat av sådana djur användes acetyl-CoA, men inte malonyl-CoA, långsamt för syntes av fettsyror. Detta ledde till antagandet att den hastighetsbegränsande reaktionen av processen som helhet är associerad med aktiviteten av acetyl-CoA-karboxylas. Faktum är att F. Linen visade att långkedjiga acylderivat av CoA vid en koncentration av 10-7 M hämmade aktiviteten av detta karboxylas. Således har själva ackumuleringen av fettsyror en hämmande effekt på deras biosyntes genom en återkopplingsmekanism.
En annan reglerande faktor i syntesen av fettsyror är tydligen citronsyra (citrat). Verkningsmekanismen för citrat är också associerad med dess effekt på acetyl-CoA-karboxylas. I frånvaro av citrat är acetyl-CoA - leverkarboxylas i form av en inaktiv monomer med en mol. väger 540 000. I närvaro av citrat förvandlas enzymet till en aktiv trimer med en mol. vikt ca. 1 800 000 och ger en 15-16-faldig ökning av synteshastigheten av fettsyror. Det kan därför antas att innehållet av citrat i levercellernas cytoplasma har en reglerande effekt på synteshastigheten av fettsyror. Slutligen, det är viktigt för syntesen av fettsyror koncentrationen av NADPH 2 i cellen.
Metabolism av omättade fettsyror
Övertygande bevis har erhållits för att i levern hos djur kan stearinsyra omvandlas till oljesyra och palmitinsyra till palmitooleinsyra. Dessa transformationer, som sker i cellmikrosomer, kräver närvaro av molekylärt syre, ett reducerat system av pyridinnukleotider och cytokrom b5. Mikrosomer kan också omvandla enkelomättade föreningar till diomättade, till exempel oljesyra till 6,9-oktadekadiensyra. Tillsammans med desaturationen av fettsyror i mikrosomer sker även deras förlängning, och båda dessa processer kan kombineras och upprepas. På detta sätt bildas till exempel nervon- och 5, 8, 11-eikosatetraensyror av oljesyra.
Samtidigt har mänskliga vävnader och ett antal djur förlorat förmågan att syntetisera vissa fleromättade föreningar. Dessa inkluderar linolsyra (9,12-oktadekadiensyra), linolensyra (6,9,12-oktadekatriensyra) och arakidonföreningar (5, 8, 11, 14-eikosatetraensyra) föreningar. Dessa föreningar klassificeras som essentiella fettsyror.Djuren upplever sin långvariga frånvaro i maten tillväxthämning och karakteristiska lesioner i hud och hår utvecklas. Fall av insufficiens av essentiella fettsyror hos människor har beskrivits. Linol- och linolensyror, som innehåller två respektive tre dubbelbindningar, samt relaterade fleromättade fettsyror (arakidonsyra, etc.) kombineras konventionellt i en grupp som kallas "vitamin F".
Biol, de essentiella fettsyrornas roll blev tydligare i samband med upptäckten av en ny klass av fysiologiskt aktiva föreningar - prostaglandiner (se). Det har fastställts att arakidonsyra och, i mindre utsträckning, linolsyra är prekursorer för dessa föreningar.
Fettsyror ingår i en mängd olika lipider: glycerider, fosfatider (se), kolesterolestrar (se), sfingolipider (se) och växer (se).
Fettsyrornas huvudsakliga plastiska funktion reduceras till deras deltagande i sammansättningen av lipider i konstruktionen av biol, membran som utgör skelettet av djur- och växtceller. I biol finns membran hl. arr. estrar av följande fettsyror: stearinsyra, palmitinsyra, oljesyra, linolsyra, linolensyra, arakidonsyra och dokosahexaensyra. Omättade fettsyror av biollipider, membran kan oxideras med bildning av lipidperoxider och hydroperoxider - den så kallade. peroxidation av omättade fettsyror.
I kroppen hos djur och människor bildas lätt endast omättade fettsyror med en dubbelbindning (till exempel oljesyra). Fleromättade fettsyror bildas mycket långsammare, varav de flesta tillförs kroppen med mat (essentiella fettsyror). Det finns speciella fettdepåer, från vilka fettsyror efter hydrolys (lipolys) av fetter kan mobiliseras för att möta kroppens behov.
Det har experimentellt visat sig att att äta fett som innehåller stora mängder mättade fettsyror bidrar till utvecklingen av hyperkolesterolemi; Användning av vegetabiliska oljor som innehåller stora mängder omättade fettsyror med mat hjälper till att minska kolesterolet i blodet (se Fettomsättning).
Medicin ägnar den största uppmärksamheten åt omättade fettsyror. Det har fastställts att deras överdrivna oxidation av peroxidmekanismen kan spela en betydande roll i utvecklingen av olika patoler, tillstånd, till exempel med strålningsskador, maligna neoplasmer, vitaminbrist E, hyperoxi och koltetrakloridförgiftning. En av produkterna av peroxidation av omättade fettsyror, lipofuscin, ackumuleras i vävnader under åldrandet. En blandning av etyletrar av omättade fettsyror, bestående av oljesyra (ca 15%), linolsyra (ca 15%) och linolensyra (ca 57%), den s.k. linetol (se), används vid förebyggande och behandling av åderförkalkning (se) och externt för brännskador och strålningsskador på huden.
I kliniken används mest metoder för kvantitativ bestämning av fria (icke-förestrade) och eterbundna fettsyror Metoder för kvantitativ bestämning av esterbundna fettsyror bygger på deras omvandling till motsvarande hydroxamsyror, som , som interagerar med Fe3+-joner, bildar färgade komplexa salter.
Normalt innehåller blodplasman från 200 till 450 mg% förestrade fettsyror och från 8 till 20 mg% icke-förestrade fettsyror. En ökning av innehållet av de senare observeras vid diabetes, nefros, efter administrering av adrenalin , under fasta, och även under känslomässig stress. En minskning av innehållet av icke-förestrade fettsyror observeras vid hypotyreos, under behandling med glukokortikoider och även efter injektion av insulin.
Individuella fettsyror - se artiklar med deras namn (till exempel arachidonsyra, arakinsyra, kapronsyra, stearinsyra, etc.). Se även Fettomsättning, Lipider, Kolesterolmetabolism.
Tabell 1. NAMN OCH FORMLER PÅ NÅGRA AV DE VANLIGASTE FETTSYROR
|
Trivialt namn |
Rationellt namn |
|||||
|
Rakkedjiga mättade fettsyror (CnH2n+1COOH) |
||||||
|
Myra |
Metan |
|||||
|
Vinäger |
Ethanova |
|||||
|
Propionsyra |
Propan |
|||||
|
Oljig |
Butan |
|||||
|
Vänderot |
Pentanic |
|||||
|
Nylon |
Hexan |
|||||
|
Enantisk |
Heptan |
|||||
|
Kapryl |
Oktan |
|||||
|
Pelargon |
Nonanova |
|||||
|
Kaprinovaya |
Dean's |
|||||
|
Undecane |
||||||
|
Lauric |
Dodekan |
|||||
|
Tridekan |
||||||
|
Myristisk |
Tetradekan |
|||||
|
Pentadekan |
||||||
|
Palmitin |
Hexadekan |
|||||
|
Margarin |
Heptadekanisk |
|||||
|
Stearin |
Oktadekan |
|||||
|
Ponadekanovaya |
||||||
|
Arachinova |
Eicosan |
|||||
|
Heneicosanovaya |
||||||
|
Begenovaya |
Docosanova |
|||||
|
Lignocerisk |
Tetrakosan |
|||||
|
Kerotiniskt |
Hexakosan |
|||||
|
Montana |
Octacosan |
|||||
|
Melissanova |
Triakontan |
CH3(CH2)28COOH |
||||
|
Lacerin |
Dotriacontane |
CH3(CH2)30COOH |
||||
|
Förgrenade mättade fettsyror (CnH2n-1COOH) |
||||||
|
Tuberkulostearisk |
10-metyloktadekan |
|||||
|
Ftionisk |
3, 13, 19-trimetyl-trikosan |
|||||
|
Ogrenade enkelomättade fettsyror (CnH2n-1COOH) |
||||||
|
Croton |
||||||
|
Kaprolinsyra |
9-decen |
CH2=CH(CH2)7COOH |
||||
|
Laureloinovap |
Dis-9-dodecene |
CH3CH2CH=CH(CH2)7COOH |
||||
|
Dis-5-dodecen |
CH3(CH2)5CH=CH(CH2)3COOH |
|||||
|
Myristolisk |
Dis-9-tetradecen |
CH3(CH2)3CH=CH(CH2)7COOH |
||||
|
Palmoljesyra |
Dis-9-hexadeken |
CH3(CH2)5CH=CH(CH2)7COOH |
||||
|
Oleic |
CH3(CH2)7CH=CH(CH2)7COOH |
|||||
|
Elaidin |
CH3(CH2)7CH=CH(CH2)7COOH |
|||||
|
Petrozelinovaja |
CH3(CH2)10CH=CH(CH2)4COOH |
|||||
|
Petroselandovaya |
CH3(CH2)10CH=CH(CH2)4COOH |
|||||
|
Vaccene |
CH3(CH2)5CH=CH(CH2)9COOH |
|||||
|
Gadoleic |
Dis-9-eikosen |
CH3(CH2)9CH=CH(CH2)7COOH |
||||
|
Cetoleic |
Cis-11-dokosen |
CH3(CH2)9CH=CH(CH2)9COOH |
||||
|
Erukovaya |
Cis-13-dokosen |
CH3(CH2)7CH=CH(CH2)11COOH |
||||
|
Nervös |
Cis-15-tetrakosen |
CH3(CH2)7CH=CH(CH2)13COOH |
||||
|
Ksimenovaya |
17-hexakosen |
CH3(CH2)7CH=CH(CH2)15COOH |
||||
|
Lumekein |
21-triakonten |
CH3(CH2)7CH=CH(CH2)19COOH |
||||
|
Ogrenade fleromättade fettsyror (CnH2n-xCOOH) |
||||||
|
Linolsyra |
||||||
|
Linelaidin |
CH3(CH2)4CH=CHCH2CH=CH(CH2)7COOH |
|||||
|
Linolensyra |
||||||
|
Linolelenaidin |
CH3CH2CH=CHCH2CH=CHCH2CH=CH(CH2)7COOH |
|||||
|
alfa-eleostearisk |
||||||
|
beta-eleostearisk |
CH3(CH2)3CH=CHCH=CHCH=CH(CH2)7COOH |
|||||
|
gamma-linolen |
CH3(CH2)4CH=CHCH2CH=CHCH2CH=CH(CH2)4COOH |
|||||
|
Punicivaya |
CH3(CH2)3CH=CHCH=CHCH=CH(CH2)7COOH |
|||||
|
Homo-gamma-linolen |
Cis-8, 11, 14, 17-eikosatrien |
CH3(CH2)7CH=CHCH2CH=CHCH2CH=CH(CH2)3COOH |
||||
|
Arachidonic |
Cis-5, 8, 11, 14-eikosatetraensyra |
CH3(CH2)4CH=CHCH2CH==CHCH2CH=CHCH2CH=CH(CH2)3COOH |
||||
|
Cis-8, 11, 14, 17-eikosatetraensyra |
CH3CH2CH=CHCH2CH=CHCH2CH=CHCH2CH=CH(CH2)6COOH |
|||||
|
Timnodonovaja |
4, 8, 12, 15, 18-eicosapen-taenoic |
CH3CH=CHCH2CH=CHCH2CH=CH(CH2)2CH=CH(CH2)2CH=CH(CH2)2COOH |
||||
|
Klupanodonovaya |
4, 8, 12, 15, 19-dokosapentaensyra |
CH3CH2CH=CH(CH2)2CH=CHCH2CH=CH(CH2)2CH=CH(CH2)2CH=CH(CH2)2COOH |
||||
|
Cis-4, 7, 10, 13, 16, 19-dokosahexaensyra |
CH3(CH2CH=CH)6(CH2)2COOH |
|||||
|
Lågland |
4, 8, 12, 15, 18, 21-tetrakosahexaen |
CH3CH2CH=CHCH2CH=CHCH2CH=CHCH2CH=CH(CH2)2CH=CH(CH2)2CH=CH(CH2)2COOH |
||||
|
Enantisk |
||||||
|
Kapryl |
||||||
|
Pelargon |
||||||
|
Kaprinovaya |
||||||
|
Undecyl |
||||||
|
Lauric |
||||||
|
Tridecyl |
||||||
|
Myristisk |
||||||
|
Pentadecyl |
||||||
|
Palmitin |
||||||
|
Margarin |
||||||
|
Stearin |
||||||
|
Icke-decyklisk |
||||||
|
Arachinova |
||||||
|
* Vid ett tryck på 100 mm Hg. Konst. |
||||||
Zinoviev A. A. Chemistry of fats, M., 1952; Newsholm E. och Start K. Regulation of metabolism, trans. från English, M., 1977; Perekalin V.V. och Sonne S.A. Organic chemistry, M., 1973; Biokemi och metodik för lipider, red. av A. R. Jonson a. J.B. Davenport, N.Y., 1971; Fettsyror, red. av K.S. Markley, punkt 1-3, N.Y.-L., 1960-1964, bibliogr.; Lipidmetabolism, red. av S. J. Wakil, N. Y.-L., 1970.
A.N. Klimov, A.I. Archakov.
Processen för fettsyraoxidation består av följande huvudsteg.
Aktivering av fettsyror. Fri fettsyra, oavsett längden på kolvätekedjan, är metaboliskt inert och kan inte genomgå några biokemiska omvandlingar, inklusive oxidation, förrän den är aktiverad. Aktivering av fettsyran sker på den yttre ytan av mitokondriella membranet med deltagande av ATP, koenzym A (HS-KoA) och Mg 2+ joner. Reaktionen katalyseras av enzymet acyl-CoA-syntetas:
Som ett resultat av reaktionen bildas acyl-CoA, som är den aktiva formen av fettsyran.
Första steget av dehydrering. Acyl-CoA i mitokondrier genomgår först enzymatisk dehydrering, och acyl-CoA förlorar 2 väteatomer i α- och β-positionerna och förvandlas till CoA-estern av en omättad syra.

Hydreringsstadiet. Omättad acyl-CoA (enoyl-CoA), med deltagande av enzymet enoyl-CoA-hydratas, fäster en vattenmolekyl. Som ett resultat bildas β-hydroxyacyl-CoA (eller 3-hydroxyacyl-CoA):

Andra steget av dehydrering. Den resulterande p-hydroxiacyl-CoA (3-hydroxiacyl-CoA) dehydreras sedan. Denna reaktion katalyseras av NAD+-beroende dehydrogenaser:

Tiolasreaktion. är klyvningen av 3-oxoacyl-CoA av tiolgruppen i den andra CoA-molekylen. Som ett resultat bildas en acyl-CoA förkortad med två kolatomer och ett tvåkolsfragment i form av acetyl-CoA. Denna reaktion katalyseras av acetyl-CoA acyltransferas (β-ketotiolas):

Den resulterande acetyl-CoA genomgår oxidation i trikarboxylsyracykeln, och acyl-CoA, förkortad med två kolatomer, går återigen upprepade gånger genom hela β-oxidationsvägen tills bildandet av butyryl-CoA (4-kolförening), som i tur oxideras upp till 2 molekyler acetyl-CoA.
Energi balans. Varje cykel av β-oxidation producerar en molekyl av FADH 2 och en molekyl av NADH. De sistnämnda, i processen med oxidation i andningskedjan och tillhörande fosforylering, ger: FADH 2 - 2 ATP-molekyler och NADH - 3 ATP-molekyler, d.v.s. totalt bildas 5 ATP-molekyler i en cykel. Oxidationen av palmitinsyra producerar 5 x 7 = 35 ATP-molekyler. I processen med β-oxidation av palmitinsyra bildas 8 molekyler acetyl-CoA, som var och en "brinner" i trikarboxylsyracykeln ger 12 molekyler ATP och 8 molekyler acetyl-CoA ger 12 x 8 = 96 molekyler ATP.
Således, totalt, med fullständig β-oxidation av palmitinsyra, bildas 35 + 96 = 131 ATP-molekyler. Med hänsyn till en ATP-molekyl som användes i början på bildningen av den aktiva formen av palmitinsyra (palmitoyl-CoA), kommer det totala energiutbytet för fullständig oxidation av en palmitinsyramolekyl under djurförhållanden att vara 131 – 1 = 130 ATP-molekyler.
Som redan nämnts får djurkroppen en betydande del av energin som utvinns under oxidationsprocessen från fettsyror, som bryts ned genom oxidation vid β-kolatomen.
β-Oxidation av fettsyror studerades första gången 19004 av F. Knoop. Det visade sig senare att β-oxidation endast sker i mitokondrier. Tack vare F. Linens och hans kollegors arbete (1954-1958) klargjordes de viktigaste enzymatiska processerna för fettsyraoxidation. För att hedra forskarna som upptäckte denna väg för fettsyraoxidation kallas β-oxidationsprocessen Knoop-Linen cykel.
β-oxidation- en specifik väg för fettsyrakatabolism, där 2 kolatomer separeras sekventiellt från karboxyländen av fettsyran i form av acetyl-CoA. Den metaboliska vägen - β-oxidation - heter så eftersom fettsyraoxidationsreaktioner sker vid β-kolatomen. Reaktionerna av β-oxidation och efterföljande oxidation av acetyl-CoA i TCA-cykeln (trikarboxylsyracykeln) fungerar som en av de viktigaste energikällorna för ATP-syntes genom mekanismen för oxidativ fosforylering. β-Oxidation av fettsyror sker endast under aeroba förhållanden.
Alla flerstegsoxidationsreaktioner accelereras av specifika enzymer. β-oxidation av högre fettsyror är en universell biokemisk process som sker i alla levande organismer. Hos däggdjur sker denna process i många vävnader, framför allt i levern, njurarna och hjärtat. Fettsyraoxidation sker i mitokondrier. Omättade högre fettsyror (oljesyra, linolsyra, linolensyra, etc.) reduceras preliminärt till mättade syror.
Inträngningen av fettsyror i mitokondriella matrisen föregås av deras aktivering genom att bilda förbindelse med koenzym A(HS~CoA), som innehåller en högenergibindning. Det senare bidrar tydligen till ett jämnare förlopp av oxidationsreaktioner av den resulterande föreningen, som kallas acylkoenzym A(acyl-CoA).
Interaktionen mellan högre fettsyror och CoA accelereras av specifika ligaser - acyl-CoA-syntetaser tre typer, specifika för syror med korta, medelstora och långa kolväteradikaler. De är lokaliserade i membranen i det endoplasmatiska retikulumet och i mitokondriernas yttre membran. Alla acyl-CoA-syntetaser verkar vara multimerer; Således har enzymet från levermikrosomer en molekylvikt på 168 kDa och består av 6 identiska subenheter. Aktiveringsreaktionen av fettsyror sker i två steg:
a) först reagerar fettsyran med ATP för att bilda acyladenylat:
RCOOH + ATP → RCO~AMP + FF
b) då sker bildningen av den aktiverade formen av acyl-CoA:
RCO~AMФ + NS~KoA → RCO~SKoA + AMF
Pyrofosfat (PP) hydrolyseras snabbt av pyrofosfatas, som ett resultat av vilket hela reaktionen är irreversibel: PP + H 2 O → 2P
Sammanfattningsekvation:
RCOOH + ATP+ HS~CoA→ RCO~SKoA + AMF + 2P
Fettsyror med kort och medelkedjelängd (från 4 till 12 kolatomer) kan tränga in i mitokondriematrisen genom diffusion, där deras aktivering sker. Långkedjiga fettsyror, som dominerar i människokroppen (12 till 20 kolatomer), aktiveras av acyl-CoA-syntetaser som finns på mitokondriernas yttre membran.
Det inre mitokondriella membranet är ogenomträngligt för långkedjiga acyl-CoA som bildas i cytoplasman. Fungerar som bärare av aktiverade fettsyror karnitin (vitamin B t), som kommer från mat eller syntetiseras från de essentiella aminosyrorna lysin och metionin.
Det yttre membranet av mitokondrier innehåller enzym karnitinacyltransferas I(karnitin palmitoyltransferas I), som katalyserar reaktionen med bildning av acylkarnitin:
RCO~SKoA + H 3 C- N + -CH 2 -CH-CH 2 -COOH ↔ H 3 C- N + -CH 2 -CH-CH 2 -COOH + HS~KoA
Acyl-CoA Karnitin (B t) Acylkarnitin Koenzym A
Detta enzym är reglerande; det reglerar hastigheten för acylgruppers inträde i mitokondrierna, och följaktligen hastigheten för fettsyraoxidation.
Det resulterande acylkarnitinet passerar genom intermembranutrymmet till den yttre sidan av det inre membranet och transporteras av karnitin acylkarnitin translokas till den inre ytan av det inre mitokondriella membranet, där enzymet karnitinacyltransferas II katalyserar överföringen av acyl till intramitokondriell CoA, det vill säga den omvända reaktionen (fig. 9).

Fig. 9. Överföring av fettsyror med långa kolväteradikaler över mitokondriella membran
Således blir acyl-CoA tillgänglig för β-oxidationsenzymer. Fritt karnitin återförs till den cytosoliska sidan av det inre mitokondriella membranet av samma translokas. Efter detta ingår acyl-CoA i β-oxidationsreaktioner.
I mitokondriematrisen sker katabolism (nedbrytning) av acyl-CoA som ett resultat av en upprepad sekvens av fyra reaktioner.
1) Den första reaktionen i varje cykel är dess oxidation av enzymet acyl-CoA-dehydrogenas, vars koenzym är FAD. Dehydrering sker mellan β- och α-kolatomerna, vilket resulterar i bildandet av en dubbelbindning i kolkedjan och produkten av denna reaktion är enoyl-CoA:
R-CH2-CH2CO~SKoA + FAD → R-CH=CHCO~SKoA + FADN2
Acyl-CoA Enoil-CoA
2) I det andra steget av fettsyraoxidationscykeln hydratiseras enoyl-CoA-dubbelbindningen, vilket resulterar i bildandet av β-hydroxiacyl-CoA. Reaktionen katalyseras av ett enzym enoyl-CoA-hydratas:
R-CH=CHCO~SKoA +H2O → R-CH-CH2CO~SKoA
Enoyl-CoA p-hydroxiacyl-CoA
3) I det tredje steget av cykeln genomgår β-hydroxyacyl-CoA dehydrering (andra oxidation) med deltagande av enzymet β-hydroxiacyl-CoA-dehydrogenas, vars koenzym är NAD+. Produkten av denna reaktion är β-ketoacyl-CoA:
R-CH-CH2CO~SKoA + NAD + → R-CОCH2CO~SKoA + NADH + H +
β-hydroxiacyl-CoA β-ketoacyl-CoA
4) Den slutliga reaktionen av fettsyraoxidationscykeln katalyseras av acetyl-CoA acyltransferas (tiolas). I detta skede reagerar β-ketoacyl-CoA med fritt CoA och klyvs för att för det första bilda ett tvåkolsfragment som innehåller de två terminala kolatomerna i moderfettsyran i form av acetyl-CoA, och för det andra ett CoA fettsyraester, nu förkortad med två kolatomer. I analogi med hydrolys kallas denna reaktion tiolys:
R-COCH 2 CO~SKoA + HS~KoA → CH 3 CO~SKoA + R 1 CO~SKoA
β-ketoacyl-CoA Acetyl-CoA Acyl-CoA,
förkortas med
2 kolatomer
Den förkortade acyl-CoA genomgår sedan nästa oxidationscykel, som börjar med en reaktion katalyserad av acyl-CoA-dehydrogenas (oxidation), följt av en hydratiseringsreaktion, en andra oxidationsreaktion, en tiolasreaktion, det vill säga denna process upprepas många gånger (Fig. 10).
β- Oxidation av högre fettsyror sker i mitokondrier. Enzymer i andningscykeln är också lokaliserade i dem, vilket leder till överföring av väteatomer och elektroner till syre under förhållanden med oxidativ fosforylering av ADP, därför är β-oxidation av högre fettsyror en energikälla för syntesen av ATP.

Fig. 10. Fettsyraoxidation
Slutprodukten av β-oxidation av högre fettsyror med jämnt antal kolatomerär acetyl-CoA, A med udda- propionyl-CoA.
Om acetyl-CoA ackumulerats i kroppen, då skulle reserverna av HS~KoA snart vara uttömda, och oxidationen av högre fettsyror skulle upphöra. Men detta händer inte, eftersom CoA snabbt frigörs från acetyl-CoA. Ett antal processer leder till detta: acetyl-CoA ingår i cykeln av trikarboxyl- och dikarboxylsyror eller glyoxylcykeln, som ligger mycket nära den, eller acetyl-CoA används för syntes av steroler och föreningar som innehåller isoprenoidgrupper, etc.
Propionyl-CoA, som är slutprodukten av β-oxidation av högre fettsyror med ett udda antal kolatomer, omvandlas till succinyl-CoA, som utnyttjas genom cykeln av trikarboxyl- och dikarboxylsyror.
Ungefär hälften av fettsyrorna i människokroppen omättad .
β-oxidation av dessa syror fortskrider på vanligt sätt tills dubbelbindningen är mellan den tredje och fjärde kolatomen. Sedan enzymet enoyl-CoA-isomeras flyttar dubbelbindningen från position 3-4 till position 2-3 och ändrar dubbelbindningens cis till transkonformation, vilket krävs för β-oxidation. I denna β-oxidationscykel inträffar inte den första dehydreringsreaktionen, eftersom dubbelbindningen i fettsyraradikalen redan är närvarande. Vidare fortsätter β-oxidationscykler, inte annorlunda än den vanliga vägen. De huvudsakliga vägarna för fettsyrametabolism visas i figur 11.

Fig. 11. Huvudvägar för fettsyrametabolism
Det upptäcktes nyligen att förutom β-oxidation, den huvudsakliga vägen för fettsyrakatabolism, hjärnvävnad α-oxidation av fettsyror med antalet kolatomer (C13-C18), det vill säga sekventiell eliminering av enkolsfragment från karboxyländen av molekylen.
Denna typ av oxidation är vanligast i växtvävnader, men kan även förekomma i vissa djurvävnader. α-Oxidation är cyklisk till sin natur, och cykeln består av två reaktioner.
Den första reaktionen består av oxidation av en fettsyra med väteperoxid till motsvarande aldehyd och CO 2 med deltagande av en specifik peroxidaser:

Som ett resultat av denna reaktion förkortas kolvätekedjan med en kolatom.
Kärnan i den andra reaktionen är hydratiseringen och oxidationen av den resulterande aldehyden till motsvarande karboxylsyra under påverkan av aldehyddehydrogenas som innehåller den oxiderade formen av koenzymet NAD:
α-oxidationscykeln upprepas sedan igen. Jämfört med β-oxidation är denna typ av oxidation energimässigt mindre gynnsam.
ω-Oxidation av fettsyror. I levern hos djur och vissa mikroorganismer finns ett enzymsystem som ger ω-oxidation av fettsyror, det vill säga oxidation vid den terminala CH 3-gruppen, betecknad med bokstaven ω. Först under påverkan monooxygenaser hydroxylering sker för att bilda ω-hydroxisyra:

ω-hydroxisyran oxideras sedan till ω-dikarboxylsyra genom verkan av motsvarande dehydrogenaser:
Den sålunda erhållna ω-dikarboxylsyran förkortas i vardera änden av β-oxidationsreaktioner.
- I kontakt med 0
- Google+ 0
- OK 0
- Facebook 0