Platelets- formed elements of blood, the amount of which is 150-400 109 / l. These are nuclear-free, pigment-free round structures that look like disks with a diameter of about 3.6 microns. They are formed in the bone marrow from large megakaryocyte cells by fragmentation of the cytoplasm; their number in the blood is constant. However, with intensive use, the rate of formation of new platelets can increase 8 times. Stimulation of thrombocytopoiesis causes thrombopoietin, which is produced in the liver and partly in the kidneys. Activation of thrombocytopoiesis can also be carried out by other hematopoietic factors, in particular interleukins (1/1-3, IL-6, IL-11), but this process is not specific compared to thrombopoietin.
Structure and function of platelets
Dense granules (β) contain non-protein substances: ADP and serotonin; factors promoting platelet aggregation, as well as antiplatelet ATP and Ca2. Lysosomal granules contain hydrolytic enzymes, and peroxisomes contain catalase. The outer shell of platelets and the VCS are coated with glycoproteins that promote platelet adhesion and aggregation.
The platelet membrane contains receptors for physiological platelet activators (ATP, adrenaline, serotonin, thromboxane Aj).
Functions of platelets:
■ platelets quickly trigger the hemostatic system. Due to adhesion (sticking) and aggregation (clumping) of platelets, a white thrombus is formed in the vessels of the microvasculature
■ substances that constrict blood vessels are released locally in the damaged area;
■ activate the onset of coagulation hemostasis with the formation of a fibrin thrombus,
■ regulate local inflammatory reactions.
In the resting state, platelets have a cytoplasmic membrane that is invaginated in places and connects to a network of channels called the open canalicular system (OCS) within the platelets. The second system of the inner membrane (dense tubular system) is formed from the endoplasmic reticulum of megakaryocytes and is not connected to the extracellular space. In the cytoplasm of non-activated platelets there are granules, including α-granules, dense β-granules, lysosome granules and peroxisomes (Fig. 9.20).
Most of all, the platelet contains α-granules containing various peptides involved in the mechanisms of coagulation, inflammation, immunity, repair and modulation of these processes.
RICE. 9.20.
Platelet activation occurs only when the vascular endothelium is damaged and there is contact with the subendothelial matrix, where collagen, other proteins, and von Willebrand factor (produced by the endothelium) are located. Platelet membrane receptors bind to von Willebrand factor (VWF), collagen, and other proteins, which leads to platelet activation, their adhesion, shape changes, and irreversible secretion of dense granules and α-granules. The change in platelet shape is caused by the intracellular system of contractile microfilaments, which leads to an increase in the surface of their membrane and the release of substances involved in coagulation hemostasis through its open tubules.
Fibrinogen attaches to the surface of the membrane due to a change in the state of its glycoproteins and promotes platelet aggregation. In platelets, thromboxane A2 is synthesized from arachidonic acid, released by the membrane of the dense tubular system, and platelet-activating factor (TAF) is synthesized, which enhances platelet aggregation and activates neutrophils. Thrombin formation also enhances platelet aggregation.
It is known that platelets synthesize blood coagulation factors V, VIII, XIII, von Willebrand factor and fibrinogen deposited in α-granules, which are released through exocytosis.
Platelet membrane lipoproteins catalyze several factors in the formation of prothrombinase. Activated platelets bind thrombin and thrombomodulin (a component of α-granules), which promotes the activation of the anticoagulant protein C.
Platelets release growth factors from α-granules into the damaged area, promote fibroblast proliferation and repair of damaged tissue. They have a connection with the humoral immune system and bind IgG, which endocytosis enters the cell, is stored in a-granules, and is then secreted by exocytosis.

RICE. 9.21. Sequence of stages of vascular-platelet homeostasis. VWF - von Willebrand factor, PF-6 - thrombostenin
Human platelets are nuclear-free, highly differentiated and highly specialized cells with a unique structure and functions.
The manifestation of functional activity by platelets is accompanied by a cardinal change in their internal structure Therefore, when studying the morphology of platelets, it is customary to distinguish cells of the “resting” stage (initial non-activated platelets) and cells that are at different stages of activation.
Resting stage platelets are described as small disc-shaped cells with a diameter of 2-5 microns. The discoid shape of platelets can be clearly observed in unfixed preparations using light microscopy. On fixed preparations stained according to Romanovsky, platelets have the appearance of polygonal, less often oval, plates, in which the peripheral part is revealed – hyalomer, and the central part – granulometer containing granules.
Normally, the hyalomer is basophilic in color, while the granulomer is oxyphilic. At the ultrastructural level, the hyalomere contains cytoskeletal elements - microtubules and actin-myosin complexes, which determine the shape of platelets at rest and during activation. The granulomere contains very small mitochondria with 1-2 cristae, glycogen accumulations, 2 types of membrane systems (open tubular system and dense tubular system), a number of lysosomes and peroxisomes, as well as secretory vesicles or granules. Elements of the vacuolar system involved in the synthesis and maturation of proteins (granular endoplasmic reticulum and Golgi apparatus) are not present in platelets or are present in the form of small residual forms that are detected only in certain pathologies. It is believed that all secretory platelet proteins are synthesized at the megakaryocyte stage .
The open tubular system (OCS) is a network consisting of single-membrane tubules and tunnels that penetrate a significant portion of the platelet volume and have contact with the plasma membrane. The membranes of CSCs include many receptor proteins and adhesion molecules; upon activation of a platelet, diffusion of these proteins from CSCs towards the plasma membrane is observed, and various membrane components - into reverse direction, That. OSC carries out the redistribution of membrane components inside platelets. In addition, CSC is involved in the exocytosis of secretory vesicles and, apparently, in the endocytosis of certain blood plasma proteins (fibronectin, albumins, immunoglobulins).
Unlike CSCs, the dense tubule system (DTS) has no connection with the plasma membrane of platelets and is a derivative of the smooth endoplasmic reticulum. The main function of PSC is the storage of intracellular calcium, which also plays an important role in the processes of platelet activation. Platelets contain a large number of secretory vesicles (vesicles), with a diameter of 200 to 600 nm; On histological preparations, these vesicles have the appearance of granules; therefore, the term “platelet granules” or “platelet granules” is most often used in the literature.
There are 3 types of granules in platelets:
1. Alpha granules– contain platelet factor IV, beta-thromboglobulin, thrombospondin, fibronectin, fibrinogen, von Willebrand factor, various growth factors (VEGF, PDGF, EGF, etc.), as well as lysosomal enzymes. Diameter of alpha granules – 300-500 nm;16
2. Beta granules(another name is dense granules) - contain ADP (non-metabolic pool), GDP, serotonin and calcium ions. Beta granules are slightly smaller than alpha granules, their diameter is 250-350 nm;
3.Gamma granules(lysosomes) - contain acid phosphatase, p-glucuronidase, cathepsin and other lysosomal enzymes. The smallest granules, their diameter is 200-250 nm.
Figure 1. Scheme of the structure of a platelet (Bykov V.L. Private human histology. St. Petersburg: Sotis, 1999. 301 p.) Mass spectrometric analysis has shown that platelets contain more than 700 types of proteins, of which about 200 have been identified to date. Most of platelet proteins are stored in alpha granules, dense bodies and lysosomes. They enter there both during megakaryocytopoiesis and by inclusion from plasma. During platelet activation, the contents of the granules are thrown out, after which the activation process becomes irreversible. It is believed that platelet degranulation is a necessary condition for their further aggregation, therefore, a violation of the functional activity of platelets is very often associated with a lack of degranulation.
Table 1 describes the chemical composition of the contents of platelet granules.
Table 1
Morphological study of the fraction of dense granules, the fraction of α-granules, the fraction of lysosomes and vesicles of human platelets allows us to adequately assess their morphofunctional status.
Blood plates
Blood platelets, or platelets, in fresh human blood they look like small, colorless, round or spindle-shaped bodies. They may aggregate (agglutinate) into small or large groups. Their number ranges from 200 to 400 x 10 9 in 1 liter of blood. Blood plates are nuclear-free fragments of the cytoplasm separated from megakaryocytes- giant bone marrow cells.
Platelets in the bloodstream are shaped like a biconvex disc. They reveal a lighter peripheral part - hyalomere and the darker, grainy part - granulometer. The platelet population contains both younger and more differentiated and aging forms. The hyalomere in young plates is colored blue (basophilene), and in mature ones - pink (oxyphilene). Young forms of platelets are larger than older ones.
The platelet plasmalemma has a thick layer of glycocalyx and forms invaginations with outgoing tubules, also covered with glycocalyx. The plasmalemma contains glycoproteins that function as surface receptors involved in the processes of adhesion and aggregation of blood platelets (i.e., the processes of blood clotting, or coagulation).
The cytoskeleton in platelets is well developed and is represented by actin microfilaments and bundles of microtubules located circularly in the hyalomere and adjacent to the inner part of the plasmalemma. Elements of the cytoskeleton ensure the maintenance of the shape of blood platelets and participate in the formation of their processes. Actin filaments are involved in reducing the volume (retraction) of blood clots that form.
The blood plates have two systems of tubules and tubules. The first is an open system of channels associated, as already noted, with invaginations of the plasmalemma. Through this system, the contents of platelet granules are released into the plasma and substances are absorbed. The second is the so-called dense tubular system, which is represented by groups of tubes that are similar to the smooth endoplasmic reticulum. The dense tubular system is the site of synthesis of cycloxygenase and prostaglandins. In addition, these tubes selectively bind divalent cations and act as a reservoir of Ca2+ ions. The above substances are necessary components of the blood clotting process.
The release of Ca 2+ ions from the tubes into the cytosol is necessary to ensure the functioning of blood platelets. Enzyme cyclooxygenase metabolizes arachidonic acid to form prostaglandins and thromboxane A2, which are secreted from the plates and stimulate their aggregation during blood coagulation.
When cyclooxygenase is blocked (for example, with acetylsalicylic acid), platelet aggregation is inhibited, which is used to prevent the formation of blood clots.
Organelles, inclusions and special granules were identified in the granulometer. Organelles are represented by ribosomes, elements of the endoplasmic reticulum of the Golgi apparatus, mitochondria, lysosomes, and peroxisomes. There are inclusions of glycogen and ferritin in the form of small granules.
Special granules make up the main part of the granulomer and are available in three types.
The first type is large alpha granules. They contain various proteins and glycoproteins involved in blood clotting processes, growth factors, and lytic enzymes.
The second type of granules is delta granules containing serotonin accumulated from plasma and other biogenic amines (histamine, adrenaline), Ca2+ ions, ADP, ATP in high concentrations.
The third type of small granules is represented by lysosomes containing lysosomal enzymes, as well as microperoxisomes containing the enzyme peroxidase.
The contents of the granules are released when the plates are activated. open system channels connected to the plasmalemma.
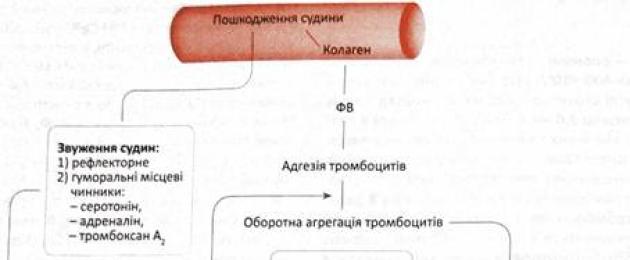
The main function of blood platelets is participation in the coagulation process, or coagulation, of blood - the body's protective reaction to damage and preventing blood loss. Platelets contain about 12 factors involved in blood clotting. When the vessel wall is damaged, the plates quickly aggregate and adhere to the resulting fibrin strands, resulting in the formation of a blood clot that closes the defect. In the process of thrombus formation, there are several stages involving many blood components.
At the first stage, platelet accumulation and the release of physiologically active substances occur. At the second stage - the actual coagulation and stopping of bleeding (hemostasis). First, active thromboplastin is formed from platelets (the so-called internal factor) and from the tissues of the vessel (the so-called external factor). Then, under the influence of thromboplastin, active thrombin is formed from inactive prothrombin. Further, under the influence of thrombin, fibrinogen is formed fibrin. Ca2+ is required for all these phases of blood coagulation.
Finally, at the last third stage, retraction of the blood clot is observed, associated with contraction of actin filaments in platelet processes and fibrin filaments.
Thus, morphologically, at the first stage, platelet adhesion occurs on the basement membrane and on the collagen fibers of the damaged vascular wall, as a result of which platelet processes are formed and granules containing thromboplastin emerge from the plates through a system of tubes onto their surface. It activates the reaction of converting prothrombin into thrombin, and the latter affects the formation of fibrin from fibrinogen.
An important function of platelets is their participation in metabolism serotonin. Platelets are practically the only blood elements in which serotonin reserves accumulate from plasma. Binding of serotonin by platelets occurs with the help of high molecular weight factors in the blood plasma and divalent cations with the participation of ATP.
During the process of blood clotting, serotonin is released from degrading platelets, which acts on vascular permeability and contraction of vascular smooth muscle cells.
The lifespan of platelets is on average 9-10 days. Aging platelets are phagocytosed by splenic macrophages. Increased destructive function of the spleen can cause a significant decrease in the number of platelets in the blood (thrombocytopenia). To correct this, the spleen may need to be removed (splenectomy).
When the number of blood platelets decreases, for example during blood loss, thrombopoietin- a factor that stimulates the formation of plates from bone marrow megakaryocytes.
· hemophilia-- hereditary disease caused by deficiency of factors VIII or IX of blood coagulation; manifested by symptoms of increased bleeding; inherited in a sex-linked recessive manner;
· purpura-- multiple small hemorrhages in the skin and mucous membranes;
· thrombocytopenic purpura-- the general name for a group of diseases characterized by thrombocytopenia and manifested by hemorrhagic syndrome (eg, Werlhof's disease);
Part four – Blood formula, leukocyte formula, age-related changes in blood, characteristics of lymph.
Hemogram and leukogram
In medical practice, blood testing plays a huge role. Clinical tests examine the chemical composition of the blood (including electrolyte composition), determine the amount of formed elements, hemoglobin, erythrocyte resistance, erythrocyte sedimentation rate and many other indicators. In a healthy person, the formed elements of blood are in certain quantitative ratios, which are usually called a hemogram, or blood formula.
The so-called differential count of leukocytes is important for characterizing the state of the body. Certain percentages of leukocytes are called leukogram, or leukocyte formula.
Age-related changes in blood
The number of red blood cells at the time of birth and in the first hours of life is higher than in an adult, and reaches 6.0-7.0 x 10 12 in 1 liter of blood. By 10-14 days it is equal to the same numbers as in an adult body. In subsequent periods, there is a decrease in the number of red blood cells with minimal indicators in the 3-6th month of life (so-called physiological anemia). The number of red blood cells returns to normal values during puberty. Newborns are characterized by the presence of anisocytosis with a predominance of macrocytes, an increased content of reticulocytes, as well as the presence of a small number of nucleated erythrocyte precursors.
The number of leukocytes in newborns is increased and reaches 30 x 10 9 in 1 liter of blood. Within 2 weeks after birth, their number drops to 9-15 x 10 9 in 1 liter (so-called physiological leukopenia). The number of leukocytes reaches a level by the age of 14-15 that remains in an adult.
The ratio of the number of neutrophils and lymphocytes in newborns is the same as in adults 4.5-9.0 x 10 9. In subsequent periods, the content of lymphocytes increases, and neutrophils decreases, and by the fourth or fifth day the number of these types of leukocytes is equalized - this is the so-called. first physiological cross leukocytes. A further increase in the number of lymphocytes and a decrease in neutrophils lead to the fact that in the 1st-2nd year of a child’s life, lymphocytes account for 65%, and neutrophils - 25%. A new decrease in the number of lymphocytes and an increase in neutrophils lead to an equalization of both indicators in 4-year-old children (this is the second physiological crossover). A gradual decrease in the content of lymphocytes and an increase in neutrophils continues until puberty, when the number of these types of leukocytes reaches the adult norm.
Lymph
Lymph is a slightly yellowish liquid tissue that flows in lymphatic capillaries and vessels. It consists of lymphoplasma (plasma lymphae) and formed elements. By chemical composition Lymphoplasm is close to blood plasma, but contains fewer proteins. Lymphoplasm also contains neutral fats, simple sugars, salts (NaCl, Na2CO3, etc.), as well as various compounds, which include calcium, magnesium, and iron.
The formed elements of lymph are represented mainly lymphocytes(98%), as well as monocytes and other types of leukocytes. Lymph is filtered from tissue fluid into blind lymphatic capillaries, where, under the influence of various factors, various components of lymphoplasma are constantly supplied from the tissues. From the capillaries, lymph moves into the peripheral lymphatic vessels, through them into the lymph nodes, then into the large lymphatic vessels and flows into the blood.
The composition of lymph is constantly changing. There are peripheral lymph (i.e. before the lymph nodes), intermediate (after passing through the lymph nodes) and central (lymph of the thoracic and right lymphatic ducts). The process of lymph formation is closely related to the flow of water and other substances from the blood into the intercellular spaces and the formation of tissue fluid.
Some terms from practical medicine:
· neonatal jaundice, physiological - transient jaundice (hyperbilirubinemia), which occurs in most healthy newborns in the first days of life;
Blood platelets, platelets, in fresh human blood look like small, colorless bodies of round, oval or spindle shape, 2-4 microns in size. They can unite (agglutinate) into small or large groups (Fig. 4.29). Their amount in human blood ranges from 2.0×10 9 /l to 4.0×10 9 /l. Blood plates are nuclear-free fragments of cytoplasm separated from megakaryocytes - giant cells of the bone marrow.
Platelets in the bloodstream are shaped like a biconvex disc. When blood smears are stained with azure-eosin, the blood platelets reveal a lighter peripheral part - the hyalomere and a darker, granular part - the granulomere, the structure and color of which can vary depending on the stage of development of the blood platelets. The platelet population contains both younger and more differentiated and aging forms. The hyalomere in young plates is colored blue (basophilene), and in mature ones – pink (oxyphilene). Young forms of platelets are larger than older ones.
In the platelet population, there are 5 main types of blood platelets:
1) young - with a blue (basophilic) hyalomere and single azurophilic granules in a reddish-violet granulomere (1-5%);
2) mature - with a slightly pink (oxyphilic) hyalomer and well-developed azurophilic granularity in the granulomere (88%);
3) old - with a darker hyalomere and granulomere (4%);
4) degenerative - with a grayish-blue hyalomere and a dense dark purple granulomere (up to 2%);
5) giant forms of irritation - with a pinkish-lilac hyalomere and violet granulomere, 4-6 microns in size (2%).
In diseases, the ratio of different forms of platelets may change, which is taken into account when making a diagnosis. An increase in the number of juvenile forms is observed in newborns. In cancer, the number of old platelets increases.
The plasmalemma has a thick layer of glycocalyx (15-20 nm), forms invaginations with outgoing tubules, also covered with glycocalyx. The plasmalemma contains glycoproteins that act as surface receptors involved in the processes of adhesion and aggregation of blood platelets.
The cytoskeleton in platelets is well developed and is represented by actin microfilaments and bundles (10-15 each) of microtubules, located circularly in the hyolomer and adjacent to the inner part of the plasmalemma (Fig. 46-48). Elements of the cytoskeleton ensure the maintenance of the shape of blood platelets and participate in the formation of their processes. Actin filaments are involved in reducing the volume (retraction) of blood clots that form.
The blood plates have two systems of tubules and tubes, clearly visible in the hyalomere under electron microscopy. The first is an open system of channels associated, as already noted, with invaginations of the plasmalemma. Through this system, the contents of platelet granules are released into the plasma and substances are absorbed. The second is the so-called dense tubular system, which is represented by groups of tubes with electron-dense amorphous material. It is similar to the smooth endoplasmic reticulum and is formed in the Golgi apparatus. The dense tubular system is the site of synthesis of cyclooxygenase and prostaglandins. In addition, these tubes selectively bind divalent cations and act as a reservoir of Ca 2+ ions. The above substances are necessary components of the blood clotting process.
| A | B | IN |
| G | D |
Rice. 4.30.Platelets. A – platelets in a peripheral blood smear. B – diagram of the structure of a platelet. B – TEM. D – non-activated (marked with an arrow) and activated (marked with two arrows) platelets, SEM. E – platelets adhered to the aortic wall in the area of damage to the endothelial layer (D, E – according to Yu.A. Rovenskikh). 1 – microtubules; 2 – mitochondria; 3 – u-granules; 4 – system of dense tubes; 5 – microfilaments; 6 – system of tubules connected to the surface; 7 – glycocalyx; 8 – dense bodies; 9 – cytoplasmic reticulum.
The release of Ca 2+ from the tubes into the cytosol is necessary to ensure the functioning of blood platelets (adhesion, aggregation, etc.).
Organelles, inclusions and special granules were identified in the granulometer. Organelles are represented by ribosomes (in young plates), elements of the endoplasmic reticulum, Golgi apparatus, mitochondria, lysosomes, and peroxisomes. There are inclusions of glycogen and ferritin in the form of small granules.
Special granules in the amount of 60-120 make up the main part of the granulomer and are represented by two main types - alpha and delta granules.
First type: a-granules- these are the largest (300-500 nm) granules, having a fine-grained central part, separated from the surrounding membrane by a small bright space. They contain various proteins and glycoproteins involved in blood clotting processes, growth factors, and hydrolytic enzymes.
The most important proteins secreted during platelet activation include lamina factor 4, p-thromboglobin, von Willebrand factor, fibrinogen, growth factors (platelet PDGF, transforming TGFp), coagulation factor - thromboplastin; Glycoproteins include fibronectin and thrombospondin, which play an important role in platelet adhesion processes. Proteins that bind heparin (thin the blood and prevent it from clotting) include factor 4 and p-thromboglobulin.
The second type of granules is δ-granules(delta granules) - represented by dense bodies 250-300 nm in size, which have an eccentrically located dense core surrounded by a membrane. There is a well-defined light space between the crypts. The main components of the granules are serotonin, accumulated from plasma, and other biogenic amines (histamine, adrenaline), Ca 2+, ADP, ATP in high concentrations.
In addition, there is a third type of small granules (200-250 nm), represented by lysosomes (sometimes called A-granules) containing lysosomal enzymes, as well as microperoxisomes containing the enzyme peroxidase. When the plates are activated, the contents of the granules are released through an open system of channels connected to the plasmalemma.
The main function of blood platelets is to participate in the process of blood clotting - the body’s protective response to damage and prevent blood loss. Platelets contain about 12 factors involved in blood clotting. When the vessel wall is damaged, the plates quickly aggregate and adhere to the resulting fibrin strands, resulting in the formation of a blood clot that closes the wound. In the process of thrombus formation, there are several stages involving many blood components.
An important function of platelets is their participation in the metabolism of serotonin. Platelets are practically the only blood elements in which serotonin reserves accumulate from plasma. Binding of serotonin by platelets occurs with the help of high-molecular factors of blood plasma and divalent cations.
During the process of blood clotting, serotonin is released from degrading platelets, which acts on vascular permeability and contraction of vascular smooth muscle cells. Serotonin and its metabolic products have antitumor and radioprotective effects. Inhibition of serotonin binding by platelets has been found in a number of blood diseases - malignant anemia, thrombocytopenic purpura, myelosis, etc.
The lifespan of platelets is on average 9-10 days. Aging platelets are phagocytosed by splenic macrophages. Increased destructive function of the spleen can cause a significant decrease in the number of platelets in the blood (thrombocytopenia). To eliminate this, surgery is required - removal of the spleen (splenectomy).
When the number of blood platelets decreases, for example during blood loss, thrombopoietin accumulates in the blood - a glycoprotein that stimulates the formation of platelets from bone marrow megakaryocytes.
Platelets and hemostasis
M.A. Panteleev1-5, A.N. Sveshnikova1-3
1Center theoretical problems Physicochemical Pharmacology RAS, Moscow; 2Federal State Budgetary Institution Federal Scientific and Clinical Center for Children's Orthopedics named after. Dmitry Rogachev, Ministry of Health of Russia, Moscow; 3Faculty of Physics, Federal State Budgetary Educational Institution of Higher Professional Education "Moscow State University them. M.V. Lomonosov";
4FGBU State Scientific Center of the Ministry of Health of Russia, Moscow; 5GemaKor LLC, Moscow
Contacts: Mikhail Alexandrovich Panteleev [email protected]
Platelets are anucleate cell fragments that play an important role in hemostasis, stopping bleeding in case of injury, as well as in pathological thrombus formation. The main way platelets perform their function is by forming aggregates that cover the site of damage. They obtain the ability to aggregate as a result of a transient process called activation. Despite their relatively simple and unambiguous function, the structure of platelets is very complex: they have an almost complete set of organelles, including the endoplasmic reticulum, mitochondria and other formations; when activated, platelets secrete a variety of granules and interact with plasma proteins and blood cells and other tissues; their activation itself is controlled by numerous receptors and complex signaling cascades. In this review, we will consider the structure of the platelet, the mechanisms of its functioning in health and disease, methods for diagnosing platelet function disorders and approaches to their correction. Particular attention will be paid to those areas of platelet science where mysteries still lurk.
Key words: platelet structure, platelet function
Platelets and hemostasis M.A. Panteleev1-5, A.N. Sveshnikova1-3
"Theoretical Problems Center of Physical and Chemical Pharmacology, Russian Academy of Sciences, Moscow; 2Federal Research Center of Pediatric Hematology, Oncology and Immunology named after Dmitriy Rogachev,
Ministry of Health of Russia, Moscow; 3Lomonosov Moscow State University, Faculty of Physics, Moscow; 4Hematological Research Center, Ministry of Health of Russia, Moscow; 5HemaCore Company, Moscow
Platelets are anuclear cell fragments playing an important role in hemostasis, termination of bleeding after damage, as well as in pathological thrombus formation. The main action of platelets is the formation of aggregates, overlapping the injury. They obtained the ability to aggregate by the transition process called activation. Despite the relatively simple and definite function platelet structure is very difficult: they have almost a full set of organelles, including the endoplasmic reticulum, mitochondria and other entities. When activated platelets secrete various granules interact with plasma proteins and red blood cells and other tissues. Their activation is controlled by multiple receptors and complex signaling cascades. In this review platelet structure, mechanisms of its functioning in health and disease, diagnostic methods of platelet function and approaches to their correction were considered. Particular attention will be given to those areas of the science of platelets, which still lay hidden mysteries.
Key words: platelet structure, platelet function
Introduction
Platelets are small, 2-4 micrometers in diameter, anucleate cellular fragments (although sometimes called cells), circulating in the bloodstream in a concentration of 200-400 thousand per microliter and are responsible for key stages of the process of stopping bleeding - hemostasis. In case of injury, they are able to attach to damaged tissues and to each other, forming a platelet plug-aggregate (Fig. 1), stopping blood loss and preventing microbes from entering the circulatory system. This is not the only mechanism of hemostasis, but it is extremely important. Hereditary and acquired disorders of platelet function, such
like Glanzmann's thrombasthenia or immune thrombocytopenia, are serious diseases characterized by dangerous bleeding. Platelets also take an active part in other components of the hemostasis mechanism: some substances they secrete cause local vasoconstriction, while others accelerate blood coagulation reactions.
On the other hand, excessive function or number of platelets, or other disorders in the cardiovascular system can lead to the formation of platelet aggregates not outside, but inside the vessel - blood clots (Fig. 2). Platelet blood clots can form in the most different situations and play a central role in such pathological conditions
Rice. 1. Hemostatic aggregate formed by platelets in the arteriole of a dog. Observed under a light microscope, a platelet plug (H) overlaps a ruptured vessel (V). The biopsy was performed 3 minutes after injury. Numerous red blood cells in the upper part of the image are located in the lumen of the wound, stretching from left to right. The size scale in the lower right corner corresponds to 10 micrometers. Reproduced from
Rice. 2. Formation of a blood clot in the arteriole. Intravital DIC microscopy of thrombus formation in a rat vessel damaged by photoactivation of pink bengal dye. A thrombus on the vascular wall covering the site of injury is indicated in the upper right part of the image. It is possible to distinguish individual platelets in it and notice that they retain their discoid shape during the first stages of attachment. The direction of flow is indicated by an arrow. The scale bar corresponds to 5 micrometers. Reproduced from
events such as heart attacks and strokes. Thus, they are responsible for the lion's share of mortality and disability in modern world, and antiplatelet drugs such as clopidogrel are among the best-selling drugs on the planet.
Platelets are simple in many ways: they have no nucleus, little or no protein synthesis, and cannot grow or divide. The platelet's task - to stick to the site of damage - also looks simple and unambiguous compared to the tasks of almost any other cell. But in practice it turns out that this simplicity is deceptive. To perform their function, they must be activated in a process that is controlled by a dozen activators acting through numerous receptors. The network of signaling pathways in the platelet that govern its response is complex and poorly understood. The platelet response itself is not a simple “sticking”, but includes dozens of functions, ranging from primary adhesion to vesiculation.
In addition to fundamental difficulties, platelets are fraught with many practical mysteries: at the moment, doctors have neither an adequate test for assessing platelet function nor an effective tool for improving it. Despite the enormous progress achieved at the end of the 20th century in connection with the development of drugs antagonistic to glycoprotein IIb-IIIa and the P2Y12 receptor, suppression of platelet activity to combat thrombosis is still not a solved problem. Finally, intensive research is now underway on the role of platelets beyond hemostasis - in angiogenesis, immunity and other systems.
Both clinical and biological research platelets attract great interest from specialists all over the world. Almost every year brings us new discoveries, and ideas about the most important processes literally within last years have undergone radical changes. In this review, we tried to focus on the fundamental concepts of the platelet and talk about the latest advances in understanding its functioning. For those who wish to become more familiar with various aspects of the life of this amazing cell, we can recommend the fundamental textbook by A.V. Mazurova. Owners English language will find valuable information in the reference textbook Platelets, edited by Alan Michelson, which is regularly reprinted.
Platelet structure
In their original, non-activated form, platelets resemble biconvex “plates” (Fig. 3, left). Due to their small size (2-4 microns in diameter), they pass freely through capillaries,
Rice. 3. Platelets. Electron micrograph of non-activated platelets retaining their discoid shape (left) and ADP-activated platelets in an aggregate (right). Reproduced from
so that their shape is constant, as opposed to the red blood cells being forced to squeeze through the capillaries. Only upon activation does the shape of the platelet change, becoming in most cases amoeboid (Fig. 3, right). The shape of the platelet is maintained both by the spectrin cytoskeleton, which gives elasticity to their shell, and by a ring of tubulin microtubules (Fig. 4), which is destroyed upon activation. The cytoplasm of the cell contains numerous granules, the main of which are dense granules containing predominantly low molecular weight substances such as serotonin and adenosine diphosphate (ADP), and alpha granules containing proteins - fibrinogen, thrombospondin, P-selectin, coagulation factor V, von Willebrand factor and many others . The contents of these granules are secreted when activated
tions. It is important to note that the platelet shape is illusory in many ways. Its internal environment is actually a continuous "sponge", a network of membrane channels that serves as an additional source of membrane surface upon activation and promotes the secretion of granules.
The ability to activate - a rapid and in most cases irreversible transition to a new state - is the main quality of a platelet. Almost any significant disturbance can serve as an activation stimulus environment, down to simple mechanical stress. However, the main physiological activators of platelets are: 1) collagen - the main protein of the extracellular matrix; 2) thrombin - serine proteinase, the central enzyme of the plasma coagulation system; 3) ADP - adenine nucleotide, which is released from destroyed vascular cells or secreted by dense granules of the platelets themselves; 4) thromboxane A2 - a lipid from the eicosanoid class, synthesized and secreted by platelets.
The action of each platelet activator is mediated through specialized receptors in the platelet membrane. Thus, collagen activates platelets through glycoprotein VI, thrombin has 2 main proteinase-activated receptors PAR1 and PAR4, ADP acts through purinoceptors P2Y1 and P2Y12. Stimulation of any of the receptors leads to the activation of a complex network of intracellular signaling cascades that control the cell's response; with different receptors generally triggering different pathways.
Membrane
Open tubular system
Microtubule ring
Dense granules
a-granules
Mitochondria
Dense tubular system
Glycogen
Dense granules
Rice. 4. Platelet structure. In the diagram on the left you can distinguish the main elements of the platelet structure, observed under an electron microscope. Reproduced from. On the right is a three-dimensional reconstruction of the insides of a platelet using electron tomography data. Note that the canalicular system shown in blue takes up a huge proportion of the cell's volume. Reproduced from
Activation of platelets is externally manifested by numerous internal rearrangements and changes in properties, the main ones among which are: 1) change in shape to amoeboid, for some platelets - spherical; 2) strengthening the ability for adhesion - attachment to the site of damage; 3) the emergence of the ability to aggregate - attach to other platelets in order to form a full-fledged plug; 4) secretion of the numerous low- and high-molecular compounds described above from dense granules, alpha granules and other sources; 5) exposure of the procoagulant membrane.
Some of these properties serve to implement the main function of platelets - the formation of a hemostatic plug, the other - to accelerate blood coagulation reactions. Thus, exposure of the procoagulant membrane and secretion of alpha granules are necessary for the implementation of the second function of platelets.
Blood clotting is a cascade of reactions in the blood plasma that ends with the formation of a network of fibrin fibers and the transfer of blood from a liquid to a jelly-like state. Many key coagulation reactions are membrane-dependent (Fig. 5), accelerating by many orders of magnitude in the presence of negatively charged phospholipid membranes, to which coagulation proteins bind through so-called calcium bridges. Under normal conditions, the platelet membrane does not support clotting reactions. Negatively charged phospholipids, primarily phosphatidylserine, are concentrated in the inner
layer of the membrane, and phosphatidylcholine in the outer layer binds coagulation factors much less well. Although some coagulation factors can bind to non-activated platelets, this does not lead to the formation of active enzymatic complexes.
Activation of the platelet presumably leads to the activation of the enzyme scramblase, which begins to quickly, specifically, bilaterally and ADP-independently transfer negatively charged phospholipids from one layer to another. As a result, an accelerated establishment of equilibrium occurs, in which the concentration of phosphatidylserine in both layers becomes equal. In addition, upon activation, exposure and/or conformational changes occur in many transmembrane proteins of the outer layer of the membrane, and they acquire the ability to specifically bind coagulation factors, accelerating reactions with their participation. Interestingly, only some platelets exhibit such properties when activated.
Generally speaking, the activated state of a platelet can be different: platelet activation has several degrees, and the expression of the procoagulant surface is one of the highest. Only thrombin or collagen can produce such a strong response. Weaker activators, especially ADP, can contribute to the functioning of strong activators. However, they are not able to independently cause the release of phosphatidylserine to the outer layer of the membrane; their effects are reduced to changing the shape, aggregation and secretion of part of the granules.
Rice. 5. Membrane blood coagulation reactions. Activation of platelets leads to the appearance of phosphatidylserine in the outer layer of the platelet membrane. Coagulation factors bind to such membranes through calcium bridges, forming protein complexes in which coagulation reactions are accelerated by orders of magnitude. The illustration shows a prothrombinase complex, consisting of factors Xa, Va, II, located on the surface of a bilayer membrane
How does a platelet work?
The most common way to test the state of the platelet hemostasis system in modern diagnostic practice is aggregation, in which the effect of adding a certain activator to a platelet suspension is assessed by optical density. An activator, most often ADP or collagen, is added to the platelet-rich blood plasma with constant stirring for several minutes. The platelets are activated, interact with each other, and the formation of aggregates occurs, which can be seen by a decrease in the turbidity of the suspension caused by a decrease in the number of light-scattering particles. There are variants of the aggregation test that involve different detection principles: for example, platelet aggregation in whole blood can be measured by using the impedance method instead of the optical method.
Perhaps, precisely in connection with the prevalence of the aggregation test over the past decades, the idea has become firmly established in the minds of many specialists that the formation of a platelet thrombus or hemostatic plug in the body occurs in a similar way: first, activation (for example, released from the cell).
current of the damaged vessel wall ADP), and then aggregation. Although the study of platelet thrombus growth in flow chambers has a history of almost half a century, only in recent decades has this traditional view begun to be questioned.
Let's consider the first stage of thrombus formation: platelet adhesion to collagen exposed at the site of damage. Let's try to estimate the times and distances typical for this process. Let the characteristic size of the damage area be, say, l = 10 micrometers (1 detached endothelial cell). Let the flow velocity be arterial, this means the flow velocity gradient on the wall is about u = 1000 s - 1. Then the platelet, which has a characteristic size (in order of magnitude) of about x = 1 micrometer, will move near the wall with a speed v = x x u = 1000 micrometers per second. This means that it will fly over the site of damage in a time of l/v = 10 microseconds, despite the fact that the typical activation time of a platelet is minutes, for some events (say, activation of integrins) several seconds, but not one hundredth of a second. This leads to the only possible conclusion, which has now been supported experimentally: in order to be activated normally, the platelet must first attach to the site of damage.
Moreover, the same applies to subsequent events of increasing thrombus size - aggregation. A platelet floating over a thrombus growing in an artery must have time to join it in hundredths of a second. Therefore, aggregation in the body can also occur only in one way: first aggregation, and then activation.
Another problem is the movement of a platelet in a vessel across the blood flow. If platelets were evenly distributed in the blood and calmly moved with a laminar flow along the vessel (and in case of injury, along the wound), each along its own flow line, then they could not approach the site of injury to perform their task in hemostasis: for adhesion the site of damage or attachment to an already activated platelet in a thrombus requires some kind of impact physical strength bringing cells into contact. In in vitro tests, this task is usually performed with a magnetic stirrer; what works in the body?
The above considerations cannot, of course, serve as evidence of a new picture of platelet hemostasis and thrombosis. This new picture, which will be outlined below, has emerged over the past 10 years as a result of active experimental work by many researchers, a leading role among which is played by the Shaun P. Jackson laboratory in Australia; Moreover, the vast majority of results were obtained using video microscopy
observations of thrombus formation in vivo. The numerical estimates presented to the reader's attention are intended only to show the unreality and internal inconsistency of the traditional idea of platelet aggregation.
How does a platelet thrombus form in reality?
The first step is the displacement of platelets to the walls of the vessel, carried out by red blood cells. Red blood cells occupy almost half of its volume; they are an order of magnitude greater than platelets in both concentration and mass. Collisions of red blood cells moving at different speeds on different flow lines lead to their redistribution and concentration near the axis of the vessel. Many details of this process are unclear, but similar redistributions have been observed in suspensions of particles of the different types, not only in the blood. Light and small platelets are constantly pushed out to the periphery, which is extremely convenient, since it is there, near potential sites of damage, that their workplace is located; Thus, the local concentration of platelets near the vessel wall is an order of magnitude higher than the average in the blood.
Moreover, even near the walls of the vessel, platelets constantly undergo collisions with red blood cells, which actually leads to the very mixing that is necessary for interaction to occur. Due to such collisions, platelets are often pressed against the wall, and if there is a site of damage there, they can attach to it. In addition to the 2 main mechanisms for which reliable theories have been built - displacement and constant pushing - others are now being discussed, but the experimental fact is indisputable: the presence of erythrocytes increases the growth rate of a platelet aggregate on a damaged surface by more than 10 times.
The second problem is the need to quickly and carefully stop a platelet that finds itself at the site of damage or near a growing blood clot. To take part in the formation of a hemostatic plug or thrombus, the platelet must extinguish its considerable speed. For this purpose, there is a special receptor on platelets, glycoprotein Ib-V-IX, and von Willebrand factor dissolved in the blood (Fig. 6). This factor, circulating in the form of large multimers up to 100 nanometers in diameter, is able to reversibly bind to collagen and platelets in the thrombus, so that it quickly coats them. Platelets rushing by cling to von Willebrand factor and begin to stop. If they were to bind collagen directly, then abruptly stopping them could be damaging, but weakly bound von Willebrand factor can detach and reattach to collagen, so that platelets can quickly recover.
to slow down, sliding just a few of its lengths, like an airplane landing on its belly.
Activation in this approach is not the first, but the last stage in the formation of a blood clot. A platelet that is reversibly bound to the site of injury may come off; however, activation can stabilize it. Platelets of the first layer, sitting directly on collagen, are activated by collagen through the glycoprotein VI receptor and then firmly bind to collagen through the integrin a2p1 receptor: proteins of this family are able to change their conformation and the strength of binding to the target under the influence of intracellular signals (Fig. 6) . In its normal state, it does not interact with collagen, but when activated, it is firmly attached to it.
The attachment of subsequent layers of platelets, i.e., the actual growth of a blood clot, occurs in a similar way: at first, the cells loosely sit on von Willebrand factor, and after activation they are securely attached through integrin receptors. The difference is that platelets communicate with each other through another integrin, called aPbp3 (or glycoprotein Pb-Sha): these receptors “grab” fibrinogen molecules from both sides and through such “fibrin-gene bridges” bind individual platelets. The second difference is that the next layers of platelets are activated not by contact with collagen (which is already covered by the first layer), but by soluble activators, which are either secreted by the platelets themselves (ADP, thromboxane A2) or formed during the operation of the plasma coagulation system (thrombin) . It is important to emphasize once again that these activators act exclusively within the thrombus: the fast flow outside it carries them away, preventing the recruitment of new cells into the thrombus.
The picture of platelet thrombus growth in vivo is now fairly well established, and the sequence of events described above is generally accepted. However, there are many unclear areas in it, which will be discussed below.
Problems in diagnosing platelet function
Currently, at least 90% of platelet function diagnostics are performed using aggregation studies. The principles and limitations of this approach have been discussed above; The main problem is that none of the aggregation tests correspond to what occurs in vivo.
Probably another 10% of the functional assessment is provided by flow cytometry, which allows us to determine the antigenic composition of proteins on the surface of the platelet. The most trained specialists can also use cytometry to characterize platelet functions in more detail: integrin activation, granule release, and phosphatidylserine. This gives useful information about individual molecules and cell abilities. However, all this does not answer the general question: how to adequately assess platelet function in general?
The most natural answer: we need to force platelets to form blood clots under conditions close to physiological. Flow chambers, in which platelet adhesion to a collagen-coated substrate is studied using microscopy, are now gaining increasing use. At the moment, commercially available cameras already exist and their standardization is underway, although to any significant extent clinical application There is still a long way to go in the practice of the diagnostic complex. A possible rival to video microscopy are similar approaches used
At GP Ib-V-IX | GP VI
non-act act
collagen
Rice. 6. The main mechanism of the initial growth of a platelet thrombus. The primary fixation of the platelet at the site of damage occurs through the interaction of the main adhesion receptor of the glycoprotein III-Y-1X with the intermediary molecule von Willebrand factor (vWF), attached to the exposed collagen (step 1). The signaling receptor glycoprotein VI then binds to collagen, leading to platelet activation (step 2). Activation of aggregation receptors of integrins a2p1 (serves for binding collagen) and aShp3 (for binding through fibrinogen bridges with other platelets) promotes the fixation of the activated platelet on collagen (step 3) and creates the basis for further growth of the thrombus. Reproduced from
in devices such as RBL, where the ability of platelets to clog with aggregates a cartridge through which whole blood is pumped is assessed.
Problems of platelet function correction
Control of platelet function is one of the main ways to combat arterial thrombosis of almost any nature. Initially, the main drug for this purpose was aspirin, which blocks the synthesis of thromboxane A2: despite the long history of the drug, only in the 2nd half of the 20th century was its ability to suppress thrombus formation and reduce the risk of heart attack discovered. In the 1990s, effective antiplatelet agents appeared that attack the fibrinogen receptor, integrin αββ3: abciximab, tirofiban, eptifibatide, as well as the domestic drug monafram. Now both of these classes of drugs are largely replaced by inhibitors of the adenosine diphosphate receptor P2Y12: these are primarily clopidogrel, as well as prasugrel, ticagrelor and cangrelor. Currently, work is actively underway to create new drugs that are more effective and have less risk of bleeding.
A more difficult task is what to do when there are few platelets or they do not work well? The technology for preparing and storing platelet concentrates for transfusion achieved its best results by the mid-1980s, and no fundamental breakthroughs have occurred since then. The short lifespan, the high risk of immune complications and infection of the patient, the continuously worsening shortage of donors throughout the world, and the absence until very recently of artificial substitutes make the situation with platelet transfusion extremely unsatisfactory, perhaps the most problematic among all blood components.
Over the course of the last decades, the only one available for clinical use
An alternative to conventional platelet concentrates was cryopreservation, which made it possible to extend their life to several years. But it was not possible to completely solve the problem of preserving the properties of platelets during freezing and thawing. In addition, freezing these cells turned out to be associated with so many technical difficulties that until now it has not been able to successfully compete with the use of unfrozen platelet concentrates.
That is why every year more and more attention was paid to the work that began back in the 1950s to create new drugs and methods that can radically extend the life and ease of use of donor platelets, or even create possible analogues that allow them to completely abandon their use. Antibacterial drugs and inhibitors of platelet activity, new cryopreservatives and freezing protocols, lyophilized platelets and vesicles based on platelet membranes, red blood cells with hemostatic function and liposomes are not a complete list of approaches used to achieve this goal. Some of them - for example, lyophilized platelets B1a$1x - are already in active clinical trials.
Mysteries of platelets
Subpopulations. One of the most intriguing mysteries of platelets is their heterogeneity. When platelets are activated, 2 subpopulations with dramatically different properties are formed. Their formation is controlled by incompletely studied signaling pathways. Interestingly, one of these subpopulations accelerates coagulation reactions, while the second is capable of normal aggregation (Fig. 7). This separation of the 2 main functions of platelets is intriguing, but no explanation has yet been offered for it.
Rice. 7. Subpopulations of blood platelets differ radically in their ability to accelerate coagulation and aggregation reactions. Dot plots of a suspension of unactivated (left) and activated (right) platelets on a flow cytometer. The x-axis shows the fluorescence of annexin V, a marker of phosphatidylserine. The y-axis shows fibrinogen fluorescence. It can be seen that upon activation, 2 subpopulations of platelets are formed, one of which is orders of magnitude higher than the other in terms of phosphatidylserine levels, but is just as inferior in fibrinogen binding. Reproduced from
Stopping the growth of a blood clot. Above we examined the sequence of events that occur during the growth of a platelet thrombus. One of the biggest problems that still remain unresolved is the question of stopping this growth: why in some cases does it go as far as complete occlusion of the vessel, while in others the vessel remains free? Now there are about a dozen hypotheses explaining the limited size of the blood clot. One of the most actively discussed is the assumption that with periodic destruction of the upper, unstable part of the thrombus, the fibrin formed inside is exposed. Nevertheless, this issue is still far from being resolved. With a high probability, there may be more than one stopping mechanism, and for different vessels these mechanisms may be different.
Platelets and the contact pathway. Quite a long time ago, researchers showed that platelets have the potential to activate blood coagulation through the contact pathway. The main candidates for the role of activators are considered to be polyphosphates released from dense granules upon activation, although there are refutations of this point of view. It appears that through this activation, the contact pathway of coagulation activation is important for platelet thrombus growth, as has been shown in recent work. This discovery allows us to hope for the creation of new antithrombotic drugs.
Microvesicles. When activated, platelets release lipid microparticles, also called microvesicles. The receptors on their surface are concentrated, and therefore these particles have enormous procoagulant activity: their surface is 50-100 times more active than the surface of activated platelets. Why platelets do this is unclear. However, in the blood of even healthy people the number of such vesicles is significant, and it significantly increases in patients with various cardiovascular and hematological diseases, correlating with the risk of thrombosis. Studying these
vesicles are hampered by their tiny size (30-300 nm), much smaller than the wavelength of light.
Platelets in oncology. Platelets play a dual role in cancer. On the one hand, they increase the risk and severity of venous thrombosis, characteristic of patients with tumors. On the other hand, they directly influence the course of the disease by regulating angiogenesis, tumor growth and metastasis through a number of mechanisms. The mechanisms of interaction between platelets and cancer cells are complex and poorly understood, but their exceptional importance is now beyond doubt.
Conclusion
Blood platelets are the most important participants in both normal hemostasis and the pathological thrombotic process, the state of which is critical for a variety of diseases and conditions. At the moment, significant progress has been made towards understanding the functioning of platelets and the correction of platelet hemostasis, but the number of scientific mysteries is still very large: the interaction of platelets with plasma hemostasis, the complexity of signaling, mechanisms for regulating the growth and arrest of platelet thrombus. Recently, information has appeared on the interaction of platelets with other body systems, indicating their important roles in immunity and morphogenesis. The main practical difficulties are the lack of adequate integral tests of platelet function and the difficulty of normalizing this function.
Acknowledgments
The authors’ work was supported by a grant from the Russian Foundation for Basic Research 14-04-00670, as well as by grants from the Basic Research Programs of the Russian Academy of Sciences “Molecular and Cellular Biology” and “ Basic Research for the development of biomedical technologies."
LITERATURE
1. Sixma J.J., van den Berg A. The haemostatic plug in haemophilia A:
a morphological study of haemostatic plug formation in bleeding time skin wounds of patients with severe haemophilia A. Br J Haematol 1984;58(4):741-53.
2. Maxwell M.J., Westein E., Nesbitt W.S.
et al. Identification of a 2-stage platelet aggregation process mediating shear-dependent thrombus formation. Blood 2007;109(2):566-76.
3. Mazurov A.V. Physiology and pathology of platelets. M.: GEOTAR-Media, 2011. 480 p.
4. Michelson A.D. Platelets. 3rd ed., 2013. London; Waltham, MA: Academic Press, xliv, 1353 p.
5. Ohlmann P., Eckly A., Freund M. et al. ADP induces partial platelet aggregation without shape change and potentiates collagen-induced aggregation in the absence of Galphaq. Blood 2000;96(6):2134-9.
6. White J.G. Electron microscopy methods for studying platelet structure and function. Methods Mol Biol 2004;272:47-63.
7.van Nispen tot Pannerden H., de Haas F., Geerts W. et al. The platelet interior revisited:
electron tomography reveals tubular alpha-granule subtypes. Blood 2010;116(7):1147-56.
8. Blair P., Flaumenhaft R. Platelet alpha-granules: basic biology and clinical correlates. Blood Rev 2009;23(4):177-89.
9. Abaeva A.A., Canault M., Kotova Y.N. et al. Procoagulant platelets form an alpha-granule protein-covered “cap” on their surface that promotes their attachment
to aggregates. J Biol Chem 2013;288(41):29621-32.
10. Kaplan Z.S., Jackson S.P. The role
of platelets in atherothrombosis. Hematology
Am Soc Hematol Educ Program 2011;2011:51-61.
11. Tanaka K.A., Key N.S., Levy J.H. Blood coagulation: hemostasis and thrombin regulation. Anesth Analg 2009;108(5): 1433-46.
12. Panteleev M.A., Ananyeva N.M., Greco N.J. et al. Two subpopulations
of thrombin-activated platelets differ in their binding of the components of the intrinsic factor X-activating complex. J Thromb Haemost 2005;3(11):2545-53.
13. Topalov N.N., Kotova Y.N., Vasil"ev S.A., Panteleev M.A. Identification of signal transduction pathways involved in the formation of platelet subpopulations upon activation. Br J Haematol 2012;157(1):105-15.
14. Yakimenko A.O., Verholomova F.Y., Kotova Y.N. et al. Identification of different proaggregatory abilities of activated platelet subpopulations. Biophys J 2012;102(10):2261-9.
15. Kotova Y.N., Ataullakhanov F.I., Panteleev M.A. Formation of coated platelets is regulated by the dense granule secretion of adenosine 5"diphosphate acting via the P2Y12 receptor. J Thromb Haemost 2008;6(9):1603-5.
16. Uijttewaal W.S., Nijhof E.J., Bronkhorst P.J. et al. Near-wall excess of platelets induced by lateral migration of erythrocytes in flowing blood. Am J Physiol 1993;264(4 Pt 2):H1239-44.
17. Tokarev A.A., Butylin A.A., Ataullakhanov F.I. Platelet adhesion from shear blood flow is controlled by near-wall rebounding collisions with erythrocytes. Biophys J 2011;100(4):799-808.
18. Turitto V.T., Weiss H.J. Red blood cells: their dual role in thrombus formation. Science 1980;207(4430):541-3.
19. Nieswandt B., Brakebusch C., Bergmeieret W. et al. Glycoprotein VI but not alpha2beta1 integrin is essential for platelet interaction with collagen. EMBO J 2001;20(9):2120-30.
20. Westein E., de Witt S., Lamers M. et al. Monitoring in vitro thrombus formation with novel microfluidic devices. Platelets 2012;23(7):501-9.
21. Favaloro E.J., Bonar R. External quality assessment/proficiency testing and internal quality control for the PFA-100 and PFA-200: an update. Semin Thromb Hemost 2014;40(2):239-53.
22. Kristensen S.D., Würtz M., Grove E.L. et al., Contemporary use of glycoprotein IIb/IIIa inhibitors. Thromb Haemost 2012;107(2):215-24.
23. Ferri N., Corsini A.,
Bellosta S. Pharmacology of the new P2Y12 receptor inhibitors: insights on pharmacokinetic and pharmacodynamic properties. Drugs 2013;73(15):1681-709.
24. Bode A.P., Fischer T.H. Lyophilized platelets: fifty years in the making. Artif Cells Blood Substit Immobil Biotechnol 2007;35(1):125-33.
25. Heemskerk J.W., Mattheij N.J., Cosemans J.M. Platelet-based coagulation: different populations, different functions.
J Thromb Haemost 2013;11(1):2-16.
26. Tosenberger A., Ataullakhanov F., Bessonov N. et al. Modeling of thrombus growth in flow with a DPD-PDE method. J Theor Biol 2013;337:30-41.
27. Bäck J., Sanchez J., Elgue G. et al. Activated human platelets induce factor XIIa-mediated contact activation. Biochem Biophys Res Commun 2010;391(1):11-7.
28. Müller F., Mutch N.J., Schenk W.A. et al. Platelet polyphosphates are proinflammatory and procoagulant mediators in vivo. Cell 2009; 139(6):1143-56.
29. Faxälv L., Boknäs N., Ström J.O. et al. Putting polyphosphates to the test: evidence against platelet-induced activation of factor XII. Blood 2013;122(23):3818-24.
30. Hagedorn I., Schmidbauer S., Pleines I. et al. Factor XIIa inhibitor recombinant human albumin Infestin-4 abolishes occlusive arterial thrombus formation without affecting bleeding. Circulation 2010;121(13):1510-7.
31. Sinauridze E.I., Kireev D.A., Popenko N.Y. et al. Platelet microparticle membranes have 50- to 100-fold higher specific procoagulant activity than activated platelets. Thromb Haemost 2007;97(3):425-34.
32. Hargett L.A., Bauer N.N. On the origin of microparticles: From “platelet dust”
to mediators of intercellular communication. Pulm Circ 2013;3(2):329-40.
33. Riedl J., Pabinger I., Ay C. Platelets in cancer and thrombosis. Hamostaseologie 2014;34(1):54-62.
34. Sharma D., Brummel-Ziedins K.E., Bouchard B.A., Holmes C.E. Platelets in tumor progression: a host factor that offers multiple potential targets in the treatment of cancer. J Cell Physiol 2014;229(8):1005-15.
- In contact with 0
- Google+ 0
- OK 0
- Facebook 0