Människans ställning i djurvärldens system
Antropologiär vetenskapen om människans ursprung.
2. Fyll i tabellen
Utveckling av åsikter om människans ursprung
3. Vad är det främsta beviset på människans ursprung från djur?
Data för jämförande anatomi och embryologi visar likheter i kroppens struktur och utvecklingen av människor och djur.
Egenskaper som är inneboende i Chordovs:
Hos människor, i de tidiga utvecklingsstadierna, representeras det inre skelettet av en ackord, neuralröret läggs på ryggsidan, kroppen har bilateral symmetri. Vidare ersätts ackordet av ryggraden, skallen bildas, fem delar av hjärnan. Hjärtat på den ventrala sidan av kroppen, det finns ett skelett av parade fria lemmar.
Egenskaper som är inneboende i klassen däggdjur:
Fem delar av ryggraden, huden är täckt med hår, det finns svett och talgkörtlar. Mata barn med mjölk, fyrkammarhjärta, varmblodighet, diafragma.
Placenta egenskaper:
Att bära ett foster i kroppen, ge fostret näring genom moderkakan.
Huvuddragen i ordningen primater:
Strukturen av lemmar, naglar, ögon i samma plan, ersättning av mjölktänder med permanenta.
Liknande egenskaper som de stora aporna. Närvaron av atavismer (drag av förfäders former som manifesterade sig i individuella individer av en given art) är en svans, multimammary, rikligt hår, etc., och rudiment (organ eller delar av kroppen som har förlorat sina ursprungliga funktioner i evolutionsprocessen och är tillgängliga hos alla individer av denna arter) är en blindtarm, visdomständer osv.
4. Beskriv människans position i den organiska världens system.
Rikets djur, Underriket Flercelliga, phylum Chordates, undertyp ryggradsdjur, klass Däggdjur, underklass Placentala, ordning Primater, underordning Apor, familj Människor (Hominider), släktet Man (Homo), art Homo sapiens (Homo sapiens), Underart Homo sapiens sapiens.
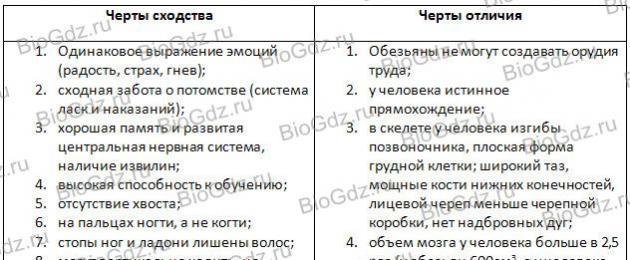
5. Gör en tabell.
Likheter och skillnader mellan människor och människoapor.

1. Definiera begreppet.
Antropogenes- en del av den biologiska evolutionen som ledde till framväxten av Homo sapiens, som separerade från andra hominider, människoapor och placenta däggdjur; processen för historisk och evolutionär bildning av den fysiska typen av en person, den initiala utvecklingen av hans arbetsaktivitet, tal.
2. Vilket koncept ligger till grund för moderna vetenskapliga idéer om människans ursprung?
I hjärtat av moderna idéer om människans ursprung är konceptet, enligt vilket människan dök upp ur djurvärlden, och de första vetenskapliga bevisen till förmån för detta koncept presenterades av Charles Darwin i hans verk The Origin of Man and Sexual Selection (1871). Forskare tror att från någon grupp av driopithecus (en utdöd grupp av högre apor), börjar den evolutionära linjen av hominider.
3. Vilka paleontologiska fynd gör det möjligt att identifiera huvudstadierna och riktningarna historisk utveckling människor och människoapor?
Analysen av paleontologiska fynd gör det möjligt att identifiera huvudstadierna och riktningarna för människans och människoapors historiska utveckling. Dessa inkluderar resterna av Australopithecus archanthropes (Pithecanthropus, Sinanthropus), Paleoanthropus (neandertalare), Cro-Magnon-mannen, deras verktyg och hällmålningar. Det vill säga att människor och moderna människoapor hade en gemensam förfader. Vidare följde deras evolutionära utveckling divergensens väg (divergens av egenskaper, ackumulering av skillnader) i samband med anpassning till specifika och olika existensvillkor.
4. Varför kan vi inte säga att människan härstammar från människoapan?
Människans och människoapornas utveckling är inte på varandra följande stadier, utan parallella grenar av evolutionen, vars diskrepans i evolutionära termer är mycket djup. Det vill säga att människor och moderna människoapor hade en gemensam förfader. Vidare följde deras evolutionära utveckling divergensens väg (divergens av egenskaper, ackumulering av skillnader) i samband med anpassning till specifika och olika existensvillkor.
5. Fyll i tabellen.
De viktigaste stadierna av antropogenes

Drivkrafter för antropogenesen
1. Definiera begreppet.
social evolution- processen med strukturell omorganisation i tiden, som ett resultat av vilket en social form eller struktur uppstår som är kvalitativt skild från den tidigare formen.
2. Rita ett diagram.
Faktorer av antropogenes.

3. Vilken roll spelar biologiska och sociala faktorer i antropogenesen?
I antropogenesen hör en viktig roll inte bara till biologiska faktorer (variabilitet, ärftlighet, urval), utan också till sociala faktorer (tal, ackumulerad erfarenhet av arbetsaktivitet och socialt beteende). Mänskliga egenskaper, på grund av sociala faktorer, fixeras inte genetiskt och ärvs inte, utan under utbildning och träning. I de första stadierna av evolutionen var urval för större anpassningsförmåga till snabbt föränderliga omständigheter av avgörande betydelse. Men senare började förmågan att överföra genetiska förvärv från generation till generation i form av en mängd olika vetenskapliga, tekniska och kulturell information spela mer och mer. viktig roll befria människan från den strikta kontrollen av naturligt urval. Sociala mönster har blivit av stor betydelse i människans evolution. Vinnarna i kampen för tillvaron var inte nödvändigtvis de starkaste, utan de som behöll de svaga: barn - befolkningens framtid, de äldre - innehavarna av information om hur man överlever. Befolkningens seger i kampen för tillvaron säkerställdes inte bara av styrka och intelligens, utan också av förmågan att offra sig i stammens namn. Människan är en social varelse, vars utmärkande drag är medvetandet, bildat på grundval av kollektivt arbete.
Sociala relationer spelar en allt större roll i utvecklingen av Homo sapiens. För moderna människor har sociala relationer och arbetsförhållanden blivit ledande och avgörande. Detta är den kvalitativa originaliteten hos mänsklig evolution.
4. Vilka moderna problem i det mänskliga samhället kan du nämna?
Människor använder alltmer surrogat och imitationer av naturlig biologisk aktivitet, upp till "virtualisering" av livet. Den mänskliga befolkningen kännetecknas av fenomen som är omöjliga för djurpopulationer. Den mänskliga befolkningen ackumulerar en genetisk börda av ärftliga sjukdomar, predisposition för sjukdomar, maligna neoplasmer, infektioner, psykiska och allergiska störningar, missanpassningsfenomen, etc. Invånare i storstäder upplever stressfenomen för överbefolkning. Många människor stödjer sin existens och kapacitet endast med hjälp av konstgjorda apparater och mediciner.
Snabb befolkningsökning skapar ekonomiska problem och förstärker människors sociala ojämlikhet. Det finns en växande klyfta mellan de maximala möjligheterna att få förmåner och deras tillgänglighet för majoriteten av människor. På olika människor livschanserna är mycket ojämlika.
Stimulerade och producerade produkter som inte bara behövs för mänskligt liv, utan också skadliga (alkohol, tobak, droger).
Alla dessa faktorer som helhet kan så småningom leda till en kris för den moderna mänskliga civilisationen, nedbrytning och försvinnande av Homo sapiens som art.
5. Vilka faktorer bidrog till utvecklingen av upprätt hållning hos människor?
Det finns ett antal hypoteser angående utvecklingen av upprätt hållning hos människor. Utseendet hos mänskliga förfäder av en S-formad ryggrad, välvd fot, förlängt bäcken, starkt korsbenet är ärftliga förändringar som bidrog till upprätt hållning.
Enligt Jan Lindblads hypotes tvingades människans apliknande förfäder att resa sig på bakbenen när de letade efter föda i vattnet och när de valde vattenhinder. Mänskliga förfäder tvingades också klättra ner från träden till marken på grund av klimat- och floraförändringar. För att leta efter mat i det höga gräset var de tvungna att resa sig från alla fyra på bakbenen.
6. Vilka fördelar och vilka problem, inklusive hälsoproblem, har en person fått i samband med bipedalism?
Individer som kunde röra sig på sina bakben var i en mer fördelaktig position. Horisonterna vidgades, händerna befriades, med hjälp av vilka mänskliga förfäder fick möjlighet att använda improviserade medel - pinnar och stenar, och därefter - att göra verktygen själva.
Nackdelarna med att gå upprätt inkluderar förlust av hastighet, en stor belastning på benen, ryggraden. Det fanns också en felaktig fördelning av blodvolymen mellan den nedre och övre halvan av kroppen. I detta fall är de nedre extremiteterna konstant i ett tillstånd av högt blodtryck, medan de övre extremiteterna är i hypotoni.
Människokroppens vertikala position leder till matsmältningsbesvär och ryggsmärtor, åderbråck och andra störningar i dess vitala funktioner. Samma orsak försvårar och försenar graviditet och förlossning extremt.
7. Kan det hävdas att den moderna människans biologiska evolution helt har stannat av?
Evolution- en kontinuerlig process, därför upphörde den inte för arten Homo sapiens. Evolutionen har flyttat till en annan nivå. En person anpassar sig till nya livsvillkor, fysiskt arbete blir mindre viktigt och utbrett. I samband med försvagningen av biologiska faktorer och ökningen av sociala faktorer kommer en person att genomgå förändringar både externa och interna, vilken exakt - det här är en tidsfråga, svaret som forskare letar efter idag.
Ursprung mänskliga raser
1. Ge definitioner av begrepp.
mänskliga raser- historiskt bildade grupperingar av människor inom arten Homo sapiens sapiens, kännetecknad av likheten mellan morfologiska och fysiologiska egenskaper.
Rasism- antivetenskaplig ideologi om ojämlikhet mellan mänskliga raser.
2. Betrakta teckningen "Race of Man". Skriv namnen på de raser du känner på den.
Australo-negroid, kaukasoid, mongoloid.
3. Vilka hypoteser om rasbildning känner du till?
Rasogenes är processen för uppkomsten och bildandet av mänskliga raser. Det finns flera hypoteser om raceogenes.
Den första är monocentrism. Forskare erkänner ett gemensamt ursprung, sociopsykologisk utveckling, en enda nivå av mental utveckling. Bevis för den monocentriska hypotesen översätter den till kategorin teorier (hypotes + bevis):
skillnader i mindre funktioner;
det finns ingen genetisk isolering;
biologiska evolutionära förändringar (minskning av skelettets massivitet, ökning av tillväxt, acceleration av utveckling) manifesteras i representanter för alla raser;
resultaten som erhölls i studien av mänskligt DNA: den första var uppdelningen av den afrikanska grenen i Negroid och Mongoloid-Caucasoid för cirka 40-100 tusen år sedan.
Polycentrister tror att raser uppstod oberoende av varandra från olika förfäder och på olika platser. Deras hypotes är mindre vetenskapligt underbyggd.
4. Vilka är de viktigaste faktorerna för rasbildning?
Naturligt urval;
mutationer;
Isolering;
blandning av befolkningar.
När människan bosatte sig på jorden, inför nya miljöförhållanden, överlevde anpassade individer och gav avkomma. De morfologiska egenskaperna som bildas under den biologiska evolutionens gång förklaras av:
karaktär miljö;
klimat;
matresurser;
mängd solljus.
5. Fyll i tabellen.
mänskliga raser

6. Vilka argument kan anföras från kritiken av rasistiska teorier?
För människor av alla raser är artens egenskaper karakteristiska:
likheten mellan kroppens struktur (strukturen av skallen, hjärnan, inre organ);
fysiologisk likhet (blodtyper, sjukdomar, försvarsreaktioner);
möjligheten till obegränsad korsning, som ett resultat av vilken fertil avkomma uppträder;
alla är av samma ursprung.
Enligt en persons gener är det omöjligt att exakt bestämma hans ras, därför finns det inga raser i genetisk mening (men bara tecken, dominerande och recessiva, olika beteendenormer). Rasskillnader är resultatet av människors anpassning till vissa existensvillkor, samt den historiska och socioekonomiska utvecklingen av det mänskliga samhället.
Homo sapiens fylogenetiska träd har endast konstruerats i allmänna termer. De viktigaste stadierna av mänsklig evolution karakteriseras i tabellen:
| Antropoider | hominider | ||||
|---|---|---|---|---|---|
| Dryopithecus | Australopithecus (Australopithecin) | skicklig man | Forntida människor (Pithecanthropus, Sinanthropus) | Forntida människor (neandertalare) | Nya människor (Cro-Magnon, mänskliga) |
| Ålder, år | |||||
| 18 miljoner | 5 miljoner | 2-3 miljoner | 2 miljoner - 200 tusen | 250-35 tusen | 50-40 tusen |
| Utseende | |||||
| Små djur med en rundad skalle, binokulärt seende, en välutvecklad hjärna; kan vara upprätt | Vikt upp till 50 kg, höjd upp till 150 cm, händerna fria, upprätt hållning | Fingrarnas falanger är tillplattade, den första tån läggs inte åt sidan | Höjd är ca 160 cm, massivt skelett, kroppsställning är halvböjd | Höjd 155-165 cm, tjocka människor, gick något böjd | Höjd är cirka 180 cm, den fysiska typen av en modern person |
| Hjärnvolym, cm 3 | |||||
| — | 550-650 | 750 | 700-1200 | Före 1400 | Runt 1400 |
| Åra | |||||
| Skallen är i struktur nära skallen hos människoapor | Massiva käkar, små framtänder och huggtänder | tänder av mänsklig typ | Skallens ben är massiva, pannan lutar, de superciliära åsarna är uttalade | Lutande panna och nackknöl, stor supraorbital kam, hakans utsprång dåligt utvecklat | Hjärnskallen dominerar över ansiktets, det finns ingen kontinuerlig supraorbital ås, hakans utsprång är väl utvecklat |
| Verktyg | |||||
| Manipulering med omgivande föremål | Systematisk användning av naturföremål | Att göra primitiva verktyg | Att göra välgjorda stenverktyg | Tillverkar olika stenverktyg | Tillverkning av komplexa verktyg och mekanismer |
| Livsstil | |||||
| flock livsstil | Flocklivsstil, jakt, samling | Kooperativ jakt och gruppskydd | Offentlig livsstil, att hålla eld, primitivt tal | Kollektiv aktivitet, omtanke om andra, utvecklat tal | riktigt tal, abstrakt tänkande, utveckling av jordbruk och industri, teknik, vetenskap, konst |
Enligt moderna paleontologiska data är människans förfäder gamla primitiva insektsätande däggdjur som gav upphov till parapithecus.
Parapithecus dök upp för cirka 35 miljoner år sedan. Dessa var trädapor, från vilka moderna gibboner, orangutanger och driopithecus härstammade.
Dryopithecus uppstod för cirka 18 miljoner år sedan. De var semi-trädlevande, halvjordiska apor som gav upphov till moderna gorillor, schimpanser och australopithecines.
australopitheciner dök upp för cirka 5 miljoner år sedan i Afrikas trädlösa stäpper. Dessa var högt utvecklade apor som rörde sig på två bakben i halvt upprätt läge. Deras höjd var 120-150 cm, kroppsvikt - 20-50 kg, hjärnvolym - cirka 600 cm 3. Med de befriade frambenen kunde de ta pinnar, stenar och andra föremål och använda dem för jakt och skydd mot fiender. Tillverkningen av verktyg av Australopithecus har inte fastställts. De levde i grupper, åt både växt- och djurmat. Australopithecus kan ha gett upphov till Homo habilis. Denna fråga är fortfarande diskutabel.
skicklig man bildades för 2-3 miljoner år sedan. Morfologiskt skiljde den sig lite från Australopithecus, men det var i detta skede som förvandlingen av apan till en man ägde rum, eftersom Handyman gjorde de första primitiva verktygen. Sedan det ögonblicket har förutsättningarna för existensen av mänskliga förfäder förändrats, som ett resultat av vilka individer med egenskaper som främjar upprätt hållning, förmågan att arbeta, förbättra de övre extremiteterna och hjärnans kognitiv aktivitet har fått fördelar i överlevnad. En skicklig man anses vara arkantropernas förfader.
Forntida människor (arkantroper)
Dessa inkluderar i synnerhet Pithecanthropus och Sinanthropus, som tillhör samma art - Homo erectus. Resterna Pithecanthropus upptäcktes 1891 på ön Java; resterna Sinanthropus- 1927 i en grotta nära Peking. Pithecanthropus och Sinanthropus var mer lika Australopithecus än moderna människor. De hade en höjd på upp till 160 cm, hjärnvolym - 700-1200 cm 3 . De levde för 2 miljoner - 200 tusen år sedan, främst i grottor och levde ett flockliv. Verktygen de gjorde var mer varierade och perfekta än Handymans. Man tror att de hade början av talet. De använde eld, vilket gjorde maten lättare att smälta, skyddade mot rovdjur och kyla och bidrog till att utöka deras utbud.
Forntida människor (paleoantroper)
De inkluderar Neandertalare. För första gången hittades deras kvarlevor i flodens dal. Neandertalare i Tyskland 1856. Neandertalare var allmänt bosatta i Europa, Afrika och Asien under istiden för 250-35 tusen år sedan. Volymen på deras hjärna nådde 1400 cm 3 . De har fortfarande superciliära åsar, en relativt låg panna, en massiv underkäke med en rudiment av ett hakutsprång. De bodde i grottor i grupper om 50-100 personer, visste hur man gör upp och underhåller eld, åt växt- och djurfoder, tillverkade olika sten-, ben- och träredskap (knivar, skrapor, yxor, pinnar etc.). De hade en arbetsfördelning: män jagade, tillverkade verktyg, kvinnor bearbetade djurkadaver, samlade in ätbara växter.
Moderna människor (neoantroper)
Neandertalarna ersattes av människor av en modern fysisk typ - cro-magnoner- de första representanterna för arten Homo sapiens. De dök upp för cirka 50-40 tusen år sedan. Under en tid existerade paleoantroper och neoantroper samtidigt, men sedan ersattes neandertalarna av Cro-Magnons. Cro-Magnons hade alla fysiska egenskaper hos levande människor: lång (upp till 180 cm), stor hjärnvolym (cirka 1400 cm 3), hög panna, utjämnade ögonbrynsryggar, utvecklat hakutsprång. Det senare indikerar ett utvecklat artikulerat tal. Cro-Magnonerna byggde bostäder, gjorde kläder av skinn sydda med bennålar, gjorde produkter av horn, ben, flinta och dekorerade dem med sniderier. Cro-Magnons lärde sig slipa, borra, kunde keramik. De levde i stamsamhällen, tämjde djur och ägnade sig åt jordbruk. De hade religionens och kulturens början.
För närvarande har vetenskapen en betydande mängd paleoantropologiska, arkeologiska och geologiska data som gör det möjligt att belysa antropogenesens förlopp (i allmänna termer). En analys av denna information ger skäl att peka ut fyra villkorade stadier (segment) av antropogenes, kännetecknade av en viss typ av fossil människa, utvecklingsnivån för materiell kultur och sociala institutioner:
1) australopithecines (människans föregångare);
2) Pithecanthropes (det äldsta folket, arkantroper);
3) Neandertalare (forntida människor, paleoantroper);
4) man modern typ, fossil och modern (neoantroper).
I enlighet med zoologisk systematik är klassificeringen av hominider som följer:
Familj - Hominidae
Underfamiljen Australopithecinae - Australopithecus
Släktet Australopithecus - Australopithecus
A. afarensis - A. afarsky A. robustus - A. kraftfull A. boisei - A boyes och andra.
Underfamiljen Homininae - människor
Genus Homo - Man
N. erectus - Man uträtad
N. sapiens neanderthalensis - neandertalarens förnuftiga man
N. sapiens sapiens - Homo sapiens rimlig.
Autralopithecines (människans föregångare)
Paleontologiska och moderna biologiska (i större utsträckning) data har bekräftat Darwins teori om människans ursprung och moderna antropologer från en gemensam initial form.
Att etablera en specifik hominoid förfader är fortfarande en utmaning för modern vetenskap. Dess existens är förknippad med en stor grupp afrikanska dryopithecus som blomstrade i miocen - pliocen (miocen sträcker sig inom 22-27 miljoner år, pliocen - inom 5-10 miljoner år), vilket leder från oligocen Egyptopithecus (30 miljoner år) . På 50-60-talet. en av driopithecus, prokonsuln, lades fram som en "modell" av den gemensamma förfadern till hominider och pongider. Den miocen dryopithecus var semi-terrestriska-semi-trädlevande apor som levde i fuktiga tropiska, bergiga eller vanliga lövskogar, såväl som skogsstäppregioner. Fynd av Miocen och Nedre Pliocen Dryopithecus är också kända i Grekland, Ungern och Georgien.
Två grenar av evolutionen avvek från den vanliga initiala formen: den första, pongid, ledde efter många miljoner år till moderna antropoida apor, den andra, hominid, till utseendet, i slutändan, av en man av den moderna anatomiska typen. Dessa två grenar utvecklades oberoende av varandra i olika adaptiva riktningar under många miljoner år. I enlighet med de naturliga och landskapsmässiga förhållandena bildades i var och en av dem specifika egenskaper hos den biologiska organisationen, motsvarande sättet att leva.
Grenen av högre apor utvecklades i riktning mot anpassning till en trädlevande livsstil, till rörelse av brachiatortyp med alla efterföljande anatomiska egenskaper: förlängning av frambenen och förkortning av bakbenen, minskning av tummen, förlängning och förträngning av bäckenet ben, utveckling av åsar på skallen, en skarp övervikt av ansiktsregionens skallar över hjärnan, etc.
Den mänskliga grenen av evolution utvecklades tvärtom i riktning mot att anpassa sig till en jordisk livsstil, upprätt gång, befria frambenen från funktionen av stöd och rörelse, använda dem för att använda naturliga föremål som verktyg, och senare till tillverkningen av konstgjorda verktyg, som var avgörande för att skilja en person från Naturlig värld. Uppfyllelsen av dessa uppgifter krävde förlängning av de nedre och förkortande av de övre extremiteterna, medan foten förlorade sina greppfunktioner och förvandlades till ett stödorgan för den uträtade kroppen, hjärnan, det huvudsakliga koordinerande hjärnorganet, utvecklades snabbt, och, följaktligen blir delen av skallen dominerande; det finns ett försvinnande av åsarna, den supraorbitala åsen, bildandet av ett hakutsprång på underkäken, etc.
Nästa viktiga fråga för evolutionär antropologi är: när uppstod en oberoende gren av mänsklig evolution och vem var dess första representant? Genom att sätta ett medelvärde för de uppskattningar som paleontologer och genetiker har erhållit får vi en period på 8-6 miljoner år. Genetiker beräknar tidpunkten för separation av de två grenarna av evolutionen baserat på de genetiska skillnaderna hos moderna hominoider och den beräknade tidpunkten för dess förekomst.
Som möjliga förfäder till hominider, förutom Ramapithecus (den senare anses ofta vara en länk i utvecklingen av orangutanger), kallas europeiska högre primater: Rudapitek och Ouranopithecus, African Kenyapithecus (en ättling till äldre prokonsuler från "driopithecus-cirkeln" ), Lufengopitek (kinesiska Ramapithecus).
Australopithecus representerar ett av de första stadierna av mänsklig evolution. De kan av de mest försiktiga utredarna betraktas som föregångare till alla fossiler och moderna människor. Australopithecus - det mest intressanta föremålet i modern mänsklig paleontologi - har blivit känt för vetenskapen sedan 30-talet av vårt århundrade. Det första fyndet av Australopithecus gjordes i södra delen av den afrikanska kontinenten. Den representerade resterna av skallen och den naturliga ebben av dess hjärndel, tillhörande ett barn.
Analysen av "ungen från Taung" visade att ett antal strukturella egenskaper skiljer sig från typen av antropoider och samtidigt liknar moderna människor. Fyndet orsakade mycket kontrovers: vissa rankade det bland de fossila antropoiderna, andra - bland de fossila hominiderna. Efterföljande fynd av sydafrikanska Australopithecus visade närvaron av två morfologiska typer - graciösa och massiva Australopithecus. Till en början tillhörde de två oberoende släkter. Flera hundra afrikanska Australopithecus är för närvarande kända. Syd- och östafrikanska massiva och graciösa varianter av Australopithecus tilldelas olika arter. Sydafrikanska arter levde i intervallet 3-1 miljon år, och östafrikanska - 4 eller mer - 1 miljon år.
Moderna antropologer tvivlar inte på att Australopithecus är en mellantyp mellan människoaporna och människan. Den huvudsakliga skillnaden från den förra är bipedal rörelse, vilket återspeglas i strukturen hos bålskelettet och vissa egenskaper hos skallen (medianpositionen för foramen magnum). Den stora bredden på bäckenbenen, förknippad med fästet av gluteal och en del av ryggradsmusklerna som rätar ut kroppen, bevisar kroppens vertikala position. En del av magmusklerna är också fästa vid bäckenskelettet, vilket stöder de inre organen när man går med en uträtad kropp.
Landskapsmiljön i Australopithecus - stäpp och skogsstäpp - krävde utveckling av förmågan att röra sig på två ben. Ibland visar antropoider denna förmåga. För Australopithecus var bipedia ett konstant inslag. Det har experimentellt bevisats att tvåfotsgång är energimässigt gynnsammare än andra typer av rörelse hos primater.
Tecken på en modern typ människa hittades på underkäkarna. Relativt små huggtänder och framtänder sticker inte ut över tändernas allmänna nivå. Ganska stora molarer har ett "mänskligt" mönster av tuberkler på tuggytan, kallat "driopithecus-mönstret". Tändernas struktur och underkäkens sammanfogning vittnar om dominansen av sidorörelser vid tuggning, vilket inte är karakteristiskt för antropoider. Australopithecus käkar är mer massiva än moderna människors. Den vertikala profilen av ansiktsregionen och dess relativt lilla totala storlek är nära den mänskliga typen. Brynet sticker fram; hjärnhålan är liten; den occipital regionen tenderar att vara rundad.
Volymen av australopithecines hjärnhålighet är liten: graciösa australopithecines - i genomsnitt 450 cm3, massiva australopithecines - 517 cm3, antropoider - 480 cm3, det vill säga nästan tre gånger mindre än för en modern person: 1450 cm3. Således är framsteg i utvecklingen av hjärnan på basis av hjärnans absoluta storlek i typen av Australopithecus praktiskt taget inte synlig. Den relativa storleken på hjärnan hos Australopithecus var i vissa fall större än antropoidernas.
Bland de sydafrikanska formerna framträder "African Australopithecus" och "Powerful Australopithecus" tydligt. Den senare kan karakteriseras enligt följande: en tjock varelse med en kroppslängd på 150-155 cm och en vikt på cirka 70 kg. Skallen är mer massiv än den afrikanska Australopithecus, underkäken är starkare. En uttalad benkrön på kronan tjänade till att fästa starka tuggmuskler. Tänderna är stora (i absolut storlek), särskilt molarerna, medan framtänderna är oproportionerligt små, så att disproportionerna mellan tänderna är tydligt synliga. Sådana morfologiska särdrag hade en vegetarisk Australopithecus, som i sin livsmiljö dras till skogens linje.
Australopithecus Africanus var mindre i storlek (graciös form): kroppslängd - upp till 120 cm och vikt - upp till 40 kg (Fig. I. 5). Att döma av kroppens ben var kroppens position vid gång mer uträtad.
Tändernas struktur motsvarade anpassning till allätande med en stor andel köttmat. Australopithecus var engagerade i insamling och jakt, möjligen med jakttroféer från andra rovdjur. Vid babianjakt använde Australopithecus stenar som kastvapen. R. Dart skapade det ursprungliga konceptet för Australopithecus-förkulturen - "osteodontokeratisk kultur", det vill säga den ständiga användningen av delar av djurskelettet som verktyg. Det föreslogs att Australopithecus mentala aktivitet blev mer komplex: detta bevisades av hög nivå deras verktygsaktivitet och utvecklade sällskapsförmåga. Förutsättningarna för dessa prestationer var bipedalism och en utvecklande hand.
Av intresse är fynden av Australopithecus och liknande former som gjorts i Östafrika, särskilt i Olduvai Gorge (Tanzania). Antropologen L. Leakey har forskat här i 40 år. Han pekade ut fem stratigrafiska skikt, vilket gjorde det möjligt att fastställa den tidsmässiga dynamiken hos de äldsta hominiderna och deras kultur under tidig Pleistocen.
Inledningsvis upptäcktes skallen av en massiv australopithecin i Olduvai Gorge, kallad "Zinjanthropus boise" ("Nötknäpparen"), senare omdöpt till "Australopithecin boise". Detta fynd är begränsat till den övre halvan av lager I (ålder 2,3-1,4 Ma). Anmärkningsvärt är de arkaiska stenredskap som finns här i form av flingor med spår av retuschering. Forskarna var förvirrade av kombinationen av stenkultur och den primitiva morfologiska typen av Australopithecus. Senare, i lager I under Zinjanthropus, hittades ben av skallen och handen från en mer avancerad människa. Det är för honom, den sk Homo habilis(Till en skicklig man) och tillhörde de gamla verktygen i Olduvai.
När det gäller Zinjanthropus (A. boisei), fortsätter den i utvecklingen av Australopithecus linjen för anpassning av massiva former till en dominerande diet av vegetabilisk mat. Denna australopithecin är större än den "kraftfulla australopithecinen" och kännetecknas av en mindre perfekt förmåga att gå på två ben (fig. I. 6).
Av stor betydelse är faktumet av samexistensen av två typer av tidiga hominider, Australopithecus Boyce och Homo habilis, bevisat av de fossila materialen i Olduvai Gorge, särskilt eftersom de skiljer sig mycket markant i morfologi och sätt att anpassa sig.
Resterna av habilis i Olduvai Gorge är inte isolerade: de samexisterar alltid med Pebble (Olduvai) kulturen, den äldsta kulturen i paleolitikum. Vissa antropologer ifrågasätter det generiska namnet

Ris. I. 6. Skalle av en supermassiv Australopithecus ("Boysova") (1,9 miljoner år)
habilis - "Nomo", föredrar att kalla honom "skicklig Australopithecus". För de flesta specialister är habilis den äldsta representanten för släktet Homo. Han använde inte bara för sina behov lämpliga föremål omgivande natur, men också modifierat dem. Antiken Homo habilis är 1,9 - 1,6 miljoner år. Fynd av denna hominid är kända i Syd- och Östafrika.
Homo habilis hade en kroppslängd på upp till 120 cm, med en vikt på upp till 40-50 kg. Käkens struktur ger ut sin förmåga att vara allätare (en egenskap hos en person). Det skiljer sig från Zinjanthropus habilis i en stor volym av hjärnhålan (volym - 660 cm3), såväl som i kranialvalvets utbuktning, särskilt i den occipitala regionen. Underkäken på habilis är mer graciös än hos andra australopitheciner, tänderna är mindre. I samband med en ganska perfekt tvåfotsgång kunde stortån röra sig, precis som hos människor, bara i vertikal riktning och foten hade valv. Habiliskroppen var nästan rak. Sålunda tog bipedia som en av antropogenesens främsta prestationer form mycket tidigt. Handen förändrades långsammare. Det finns ingen perfekt opposition av tummen till resten, dess dimensioner, att döma av benelementen, är små. Fingrarnas falanger är böjda, vilket inte är typiskt för en modern person, men de terminala falangerna är platta.
I lagren av Oluvai Gorge (ålder från 1,2-1,3 miljoner år) hittades benrester av former som kan tolkas som en övergång från typen av progressiv Australopithecus till typen av Pithecanthropus. Pithecanthropus har också upptäckts på denna plats.
Det är svårt att tolka och klassificera former som liknar Afrikas Australopithecus, men som finns utanför detta fastland. Så på ön Java upptäcktes ett fragment av underkäken hos en högre primat, vars övergripande dimensioner avsevärt översteg dimensionerna hos moderna människor och de största aporna. Han fick namnet "Meganthropus Paleo-Javanese". För närvarande hänvisas det ofta till Australopithecus-gruppen.
Alla dessa australopitheciner och tidiga representanter för släktet Homo föregicks i tiden av den graciösa "Afar Australopithecus" (A. afarensis), vars benrester upptäcktes i Etiopien och Tanzania. Antiken för representanter för denna art är 3,9-3,0 miljoner år. Den lyckliga upptäckten av ett mycket komplett skelett av ämnet, som heter "Lucy", tillåter oss att representera Afar australopithecines enligt följande. Kroppsmåtten är mycket små: kroppslängd - 105-107 cm, vikten översteg något över 29 kg. I strukturen av skallen, käkarna och tänderna noterades mycket primitiva tecken. Skelettet är anpassat till en tvåfotad gång, även om det skiljer sig från en mänsklig. Studien av fotspår i vulkanisk aska (antik - minst 3,6 miljoner år) leder till slutsatsen att Afar australopithecinerna inte sträckte ut sina ben helt vid höftleden, och när de gick korsade de sina fötter och placerade dem en framför Övrig. Foten kombinerar progressiva drag (stor och adducerad första tå, uttalad fotvalv, formad häl) och apliknande drag (dansen är inte orörlig). Proportionerna av den övre
och nedre extremiteter motsvarar upprätt hållning, men det finns tydliga tecken på anpassning till det trädlevande rörelsesättet. I handen kombineras även progressiva tecken med arkaiska (relativ förkortning av fingrarna) förknippade med förmågan till trädgående rörelse. Tecken på "force capture" som är karakteristiska för hominider observeras inte. Som primitiva egenskaper hos skallen bör ett starkt utsprång av ansiktsregionen och en utvecklad occipital lättnad noteras. Utskjutande huggtänder och diastemas mellan tänderna i över- och underkäken ser arkaiska ut även mot bakgrund av andra Australopithecus. Molarerna är mycket stora och massiva. Den absoluta storleken på hjärnan hos Afar Australopithecus går inte att skilja från storleken på antropomorfa apor, men dess relativa storlek är något större. Individuella Afar-individer har en tydlig "schimpansisk" morfologi, vilket bevisar en inte så avlägsen separation av de evolutionära grenarna av hominider och pongider.
Vissa neurologer tror att i mycket gamla representanter för Australopithecus är det redan möjligt att fixa den strukturella omstruktureringen av de parietala, occipitala och temporala regionerna i hjärnan; samtidigt är bland annat hjärnans yttre morfologi omöjlig att skilja från en apa. Hjärnomstrukturering kan börja på cellnivå.
De mest moderna paleoantropologiska upptäckterna gör det möjligt att preliminärt identifiera arten av Australopithecus, som föregick "afarianerna" i tid. Dessa är den östafrikanska Australopithecus A. ramidus (Etiopien) (representerad av underkäken) och A. anamensis (Kenya); (representerad av fragment av tuggapparaten). Forntiden för båda fynden är cirka 4 miljoner år. Det finns också äldre fynd av australopitheciner som inte har en artdefinition. De fyller det tillfälliga uppehållet mellan de äldsta australopithecinerna och den hominoida förfadern.
Av stort intresse är fynden av tidiga företrädare för släktet Homo, som gjordes på sjöns östra strand. Turkana (Kenya). Progressiva tecken på Homo habilis "1470" inkluderar en hjärnvolym på cirka 770 cm3 och en utjämnad lättnad av skallen; antiken - cirka 1,9 miljoner år.
Vilken plats tog verktygsaktiviteten i Australopithecus evolutionära prestationer? Antropologer har inte en enig åsikt om det oupplösliga i sambandet mellan redskapsaktivitet och tvåfotsgång. Trots fynden av mycket uråldriga stenverktygskulturer finns det en betydande tidsskillnad mellan framväxten av tvåfoting och uppkomsten av arbetskraft. Det antas att orsaken till isoleringen av de första hominiderna från djurvärlden kan vara överföringen av den defensiva funktionen hos dentalapparaten till konstgjorda försvarsverktyg, och användningen av verktyg blev en effektiv anpassning i de första människornas beteende som bosatte savannen. Monument av Olduvai-kulturen klargjorde inte frågan om kopplingen mellan Australopithecus och Olduvai-verktyg. Således är det känt att hitta benen av den progressiva "habilis" och den massiva Australopithecus i samma horisont med Olduvai-verktygen.
De äldsta verktygen hittades i äldre horisonter än fragmenten av de första obestridliga företrädarna för släktet Homo. De paleolitiska kulturerna i Kenya och Etiopien är alltså 2,5-2,6 miljoner år gamla. Analysen av nya material visar att Australopithecus bara kunde använda verktyg, men att endast representanter för släktet Homo kunde göra dem.
Olduvai (sten) eran är den tidigaste under paleolitikum (gammal stenålder). De mest karakteristiska verktygen är massiva arkaiska artefakter gjorda av småsten och fragment av sten, såväl som stenar - ämnen (kärnor), verktyg på flingor. Ett typiskt Olduvai-verktyg är en hackare. Det var en sten med fasad ände, vars obearbetade del tjänade till att hålla redskapet i handen (fig. I. 7). Bladet kunde bearbetas på båda sidor; redskap med flera facetter och bara slagstenar hittades också. Olduvai-verktyg skiljer sig i form och storlek, men har samma typ av blad. Detta beror på syftet med åtgärder för att utveckla verktyg. Arkeologer noterar att det redan från början av paleolitikum fanns en uppsättning verktyg för olika ändamål. Fynd av brutna ben tyder på att Australopithecus var jägare. Olduvai-verktyg överlever till sena tider, särskilt i Syd- och Sydostasien. Den långa existensen av Olduvai (1,5 miljoner år) åtföljdes nästan inte av tekniska framsteg. Australopithecus kunde ordna enkla skydd som vindbarriärer.

Ris. I. 7. Nedre paleolitikums Olduvai-kultur. Pithecanthropes
(tidigaste människor, arkantroper)
Pithecanthropes är den andra stadiala gruppen av hominider efter Australopithecus. I denna aspekt kallas de i den specialiserade litteraturen ofta (alla varianter av gruppen) som "arkantroper", d.v.s. "det äldsta folket"; här kan du också lägga till definitionen av "sanna människor", eftersom Pithecanthropus tillhörighet till familjen hominider inte ifrågasätts av någon av antropologerna. Tidigare kombinerade vissa forskare Pithecanthropus med neandertalare i ett evolutionärt stadium.
Pithecanthropusfynd är kända i tre delar av världen - Afrika, Asien och Europa. Deras förfäder var representanter för Homo habilis (senare östafrikanska representanter för denna art kallas ofta Homo rudolfensis). Tiden för existensen av pithecanthropes (inklusive de tidigaste, Homo ergaster) kan representeras i intervallet 1,8 miljoner år - mindre än 200 tusen år. De äldsta representanterna för scenen upptäcktes i Afrika (1,6 miljoner år - 1,8 miljoner år); från 1 miljon årsskiftet är de vanliga i Asien, och från tiden för 0,5 miljoner år levde pithecanthropes (ofta kallade "preneandertalare", eller representanter för Homo heidelbergensis) i Europa. Pithecanthropes nästan över hela världen kan förklaras av deras ganska höga biologiska och sociala utveckling. Utvecklingen av olika grupper av Pithecanthropes skedde i olika hastigheter, men hade en riktning - mot sapiens-typen.
För första gången upptäcktes benfragmenten från Pithecanthropus av den holländska läkaren E. Dubois ungefär. Java 1891. Det är anmärkningsvärt att fyndets författare delade konceptet om en "mellanlänk" i den mänskliga genealogin, som tillhörde darwinisten E. Haeckel. Nära byn Trinil hittades (successivt) den övre molaren, dödskallen och lårbenet. Den arkaiska karaktären hos kranialhöljet är imponerande: en sluttande panna och en kraftfull supraorbital kam och en helt modern typ av lårben. Lagren som innehåller Trinil-faunan går tillbaka till 700 tusen år sedan (för närvarande 500 tusen år). 1894 gav G. Dubois först vetenskaplig beskrivning"Pitpecanthropus erectus" ("apa-man erectus"). Vissa europeiska forskare mötte en sådan fenomenal upptäckt med misstro, och Dubois själv trodde ofta inte på dess betydelse för vetenskapen.
Med ett intervall på 40 år gjordes andra fynd av Pithecanthropes ca. Java och på andra ställen. I Pungat-lagren med Dzhetis-faunan nära byn Mojokerto upptäcktes en babyskalle av en Pithecanthropus. Fyndets ålder är nära 1 miljon år. Fynd av ben i skallen och skelettet gjordes i Sangiran-orten (uråldriga omkring 800 tusen år) under 1936-1941. Nästa fyndserie nära Sangiran avser perioden 1952-1973. Det mest intressanta fyndet var skallen av en Pithecanthropus med en bevarad ansiktsdel av skallen, gjord 1963. Resterna av en paleolitisk kultur på ungefär. Java hittades inte.
En fossil man som liknar Pithecanthropus hittades i mitten av Pleistocene avlagringar i Kina. Sinanthropus (kinesiska Pithecanthropus) tänder upptäcktes i kalkstensgrottan i Zhoukou-dian 1918. Samlingen av slumpmässiga fynd ersattes av utgrävningar och 1937 upptäcktes resterna av mer än 40 Sinanthropus-individer på denna plats (fig. 1,8). Beskrivningen av denna variant av Pithecanthropus gjordes först av den kanadensiska specialisten Vlekom. Den absoluta dateringen av Sinanthropus uppskattas till 400-500 tusen år. Benresterna av Sinanthropus åtföljs av många kulturella

rester (stenredskap, krossade och brända djurben). Av störst intresse är den flera meter tjocka aska som finns i Sinanthropus jaktläger. Användningen av eld för att bearbeta mat gjorde den mer smältbar, och det långsiktiga underhållet av en brand indikerar en ganska hög nivå av utveckling av sociala relationer mellan Sinanthropes.
Flera fynd tillåter oss att med tillförsikt tala om verkligheten för Pithecanthropus-taxonen. Här är huvuddragen i dess morfotyp. Den moderna typen av lårbenen och positionen av foramen magnum, liknande vad vi ser på moderna kranier, vittnar om Pithecanthropus otvivelaktiga anpassning till upprätt hållning. Den totala massiviteten hos Pithecanthropus-skelettet är större än Australopithecus. Många arkaiska egenskaper observeras i skallens struktur: en högt utvecklad lättnad, en sluttande frontalregion, massiva käkar, uttalad prognatism av ansiktsregionen. Skallens väggar är tjocka, underkäken är massiv och bred, tänderna är stora, medan hundens storlek är nära modern. En högt utvecklad occipital lättnad är förknippad med utvecklingen av livmoderhalsmusklerna, som spelade en betydande roll för att balansera skallen när man går. Uppskattningar av storleken på hjärnan hos Pithecanthropes som ges i modern litteratur varierar från 750 till 1350 cm3, dvs. motsvarar ungefär det minsta lägre tröskel värden som ges för australopitheciner av habilis-typ. Tidigare jämförda arter tillskrevs en signifikant skillnad. Endokranernas struktur vittnade om komplikationen av hjärnans struktur: i större utsträckning har Pithecanthropes utvecklat områden i parietalregionen, nedre frontala och övre bakre delarna av frontalregionen, som är förknippad med utvecklingen av specifika mänskliga funktioner - arbete och tal. På synantropernas endokraner hittades nya tillväxtfokus associerade med bedömning av kroppsposition, tal och fina rörelser.
Sinanthropus är något annorlunda i typ från Pithecanthropus. Längden på dess kropp var cirka 150 cm (Pithecanthropus - upp till 165-175 cm), skallens dimensioner ökade, men typen av struktur var densamma, med undantag för en försvagad occipital relief. Skelettet av Sinanthropus är mindre massivt. Anmärkningsvärt är den graciösa underkäken. Hjärnans volym är mer än 1000 cm3. Skillnaden mellan Sinanthropus och Javanese Pithecanthropus bedöms på underartnivå.
Naturen hos matrester, såväl som strukturen hos underkäkarna, indikerar en förändring i typen av utfodring av synantroper mot allätare, vilket är ett progressivt tecken. Sinanthropus har sannolikt kannibalism. I frågan om deras förmåga att göra eld var arkeologer oense.
Analys av mänskliga benrester från denna fas av antropogenesen gör att vi kan rekonstruera ålders- och könssammansättningen för synanthropusgrupper: 3-6 män, 6-10 kvinnor och 15-20 barn.
Kulturens jämförande komplexitet kräver en tillräckligt hög nivå av kommunikation och ömsesidig förståelse, därför är det möjligt att förutsäga förekomsten av primitivt tal vid denna tidpunkt. Den biologiska grunden för en sådan prognos kan betraktas som en ökning av benavlastningen på platserna för fastsättning av tungans muskler, början av bildandet av hakan och graciliseringen av underkäkarna.
Fragment av skallar från antiken, i proportion till de tidiga Pithecanthropes av Fr. Java (cirka 1 miljon år gammal), som finns i två provinser i Kina - Lantian, Kuvanlin. Det är intressant att de mer antika kinesiska Pithecanthropes skiljer sig från Sinanthropes på samma sätt som de tidiga Pithecanthropes från de senare, nämligen i den större massiviteten hos benen och den mindre storleken på hjärnan. Sen progressiv Pithecanthropus inkluderar ett nyligen hittat i Indien. Här hittades tillsammans med Late Acheulean verktyg en skalle med en volym på 1300 cm3.
Verkligheten av existensen av Pithecanthropus-stadiet i antropogenesen är praktiskt taget inte ifrågasatt. Det är sant att de senare representanterna för Pithecanthropes anses vara förfäder till efterföljande, mer progressiva former. Frågan om tid och plats för uppkomsten av den första Pithecanthropus har diskuterats mycket inom vetenskapen. Tidigare ansågs Asien vara dess hemland, och tiden för uppkomsten uppskattades till cirka 2 miljoner år. Nu är det här problemet löst på ett annat sätt. Afrika anses vara födelseplatsen för både Australopithecus och Pithecanthropus. 1984, i Kenya (Nariokotome), upptäcktes en 1,6 miljoner år gammal Pithecanthropus (komplett skelett av en tonåring). Huvudfynden av de tidigaste pithecanthropes i Afrika är: Koobi Fora (1,6 miljoner år), sydafrikanska Swartkrans (1,5 miljoner år), Olduvai (1,2 miljoner år). Afrikansk pithecanthropus av kusten Medelhavet(Ternifin) har en forntid på 700 tusen år. Den geologiska antiken för de asiatiska varianterna kan uppskattas till 1,3–0,1 Ma. Arkeologiska bevis från platser i Mellanöstern, närmare Afrika än Asien, är kända, vilket tyder på att antiken för afrikanska pithecanthropes kan nå 2 miljoner år.
De synkrona formerna av fossilmannen från Europa är yngre och ganska säregna. De kallas ofta för "pre-neandertalare" eller hänvisas till som Homo heidelbergensis, som i Afrika, Europa och Asien var förfäder för moderna människor och neandertalarna i Europa och Asien. Europeiska former har följande ålder: Mauer (500 tusen år), Arago (400 tusen år), Petralona (450 tusen år), Atapuerca (300 tusen år). Broken Hill (300 tusen år) och Bodo (600 tusen år) har en evolutionär övergångskaraktär i Afrika.
I Kaukasus är det äldsta fyndet i Georgien Dmanisi-mannen, vars antika uppskattas till 1,6-1,8 miljoner år. Anatomiska egenskaper gör det möjligt att ställa den i paritet med de äldsta hominider i Afrika och Asien! Pithecanthropes hittades också på andra platser: i Uzbekistan (Sel-Ungur), i norra Kaukasus (Kudaro), Ukraina. En form mellan Pithecanthropes och Neandertalare hittades i Azerbajdzjan (Azykh). Den Acheulean mannen bodde tydligen på Armeniens (Jerevan) territorium.
Tidiga pithecanthropes skiljer sig från senare i större benmassa och en mindre hjärnstorlek. En liknande skillnad observeras i Asien och Europa.
I paleolitikum motsvarar Acheulean den fysiska typen av Pithecanthropus och tidiga neandertalare. Ashelens ledande verktyg är en handyxa (fig. I. 9). Det visar en hög nivå i utvecklingen av stenbearbetningsteknik. Inom gränserna för den Acheuliska eran kan man observera en ökning av grundigheten hos efterbearbetning av axlar: antalet spån från verktygets yta ökar. Ytfinishen blir finare när stenflismaskiner byts ut mot mjukare av ben, horn eller trä. Storleken på en handyxa nådde 35 cm. Den var gjord av sten genom att flisa på båda sidor. Yxan hade en spetsig ände, två längsgående blad och en rå motstående egg. Man tror att yxan hade olika funktioner: den fungerade som ett slaginstrument, användes för att gräva upp rötter, stycka djurkroppar och bearbeta trä. I de södra regionerna finns en yxa (fock), som kännetecknas av ett tvärgående blad, inte korrigerat genom retuschering, och symmetriskt bearbetade kanter.

En typisk Acheulean-yxa förbrukar inte all den tekniska mångfalden som kännetecknar den perioden. Det fanns en fling-"klekton"-kultur, såväl som en progressiv flingkultur "Levallois", som kännetecknas av tillverkning av verktyg från flingor av skivformade ämnen, ytan på ämnena bearbetades preliminärt med små chips. Förutom yxor finns små verktyg som spetsar, skrapor och knivar på Acheulean-platserna. Några av dem överlever till Cro-Magnons tid. Det finns också Olduvai-verktyg i Acheulean. Sällsynta träverktyg är kända. Man tror att Asiens Pithecanthropus kunde nöja sig med bambuverktyg.
Jakt var av stor betydelse i Acheuleanernas liv. Pithecanthropes var inte bara samlare. Acheulean-monumenten tolkas som jaktläger, eftersom ben från stora djur finns i deras kulturlager. Acheulean-kollektivens liv var svårt, människor var engagerade i olika typer av arbete. Olika typer av läger är öppna: jaktläger, verkstäder för flintbrott, långtidsläger. Acheuleanerna byggde bostäder på öppna platser och i grottor. I området Nice öppnades en bosättning av hyddor.
Den naturliga miljön för den Acheulean mannen bestämde egenskaperna hos den materiella kulturen. Typerna av verktyg på olika platser finns i olika proportioner. Jakten på stora djur krävde en nära sammankomst av ett team av människor. Parkeringsplatser av olika slag vittnar om att det finns en arbetsfördelning. Resterna av härdar talar om effektiviteten av Pithecanthropes användning av eld. På den kenyanska platsen Chesovanja är spåren av eld 1,4 miljoner år gamla. Neandertalmannens Mouster-kultur är utvecklingen av de tekniska landvinningarna från Pithecanthropes änglakultur.
Som ett resultat av de första människornas afro-asiatiska magrationer uppstod två huvudcentra för mänsklig evolution - västerländskt och östligt. Pithecanthropus-populationer åtskilda av stora avstånd kunde utvecklas under lång tid isolerade från varandra. Det finns en åsikt att neandertalare inte var ett naturligt utvecklingsstadium i alla regioner, i Afrika och Europa var Pithecanthropes ("preneandertalare") sådana.
Neandertalare (forntida människor, paleoantroper)
I den traditionella stadialmodellen av antropogenes representerades det mellanliggande evolutionära steget mellan Homo erectus och Homo sapiens av paleoantroper ("forntida människor"), som i absolut kronologi levde från 300 tusen år till cirka 30 tusen år i Europa, Asien och Afrika. I icke-professionell litteratur kallas de ofta för "neandertalare", efter namnet på ett av de första fynden 1848 i Neandertalområdet (Tyskland).
I allmänhet fortsätter paleoantroper evolutionen av "Human erectus" (mer exakt, Homo heidelbergensis), men i moderna system kallas de ofta för en sidogren av hominider. När det gäller den allmänna nivån av evolutionära prestationer är dessa hominider närmast moderna människor. Därför har de genomgått förändringar i sin status i klassificeringen av hominider: paleoantroper anses för närvarande som en underart av Homo sapiens, det vill säga som dess fossila variant (Homo sapiens neanderthalensls). Denna syn återspeglar ny kunskap om komplexiteten i neandertalbiologi, intelligens och social organisation. Antropologer som lägger stor vikt vid de biologiska skillnaderna mellan neandertalare och moderna människor betraktar dem fortfarande som en separat art.
De första fynden av neandertalare gjordes på 1800-talet. i Västeuropa och hade inte en entydig tolkning.
Grupper av paleoantroper, belägna inom ett betydande intervall av geologisk tid, är mycket olika i morfologiskt utseende. Antropologen V.P. Alekseev gjorde ett försök att klassificera grupper av neandertalare, liknande morfologiskt och kronologiskt, och pekade ut flera grupper: europeiska, afrikanska, Skhul-typ och västasiatiska. De flesta fynden av paleoantroper från Europa är kända. Ofta bebodde neandertalarna glaciärzonerna.
På samma grunder (morfologiska och kronologiska), bland de europeiska formerna av den angivna tiden, urskiljs följande nivåer: "de tidigaste neandertalarna" - "pre-neandertalarna", "tidiga neandertalarna" och "sena neandertalarna".
Antropologer föreslog att det objektivt sett fanns flera övergångar mellan på varandra följande stadialgrupper, därför kunde en evolutionär övergång till paleoantroper i olika områden ha ägt rum från flera varianter av Pithecanthropus. Representanter för arten Homo heidelbergensis kan vara föregångare (Petralona, Swanscombe, Atapuerca, Arago, etc.).
Den tidigaste europeiska gruppen inkluderar den fossila skallen från Steinheim-platsen (200 tusen år gammal), som hittades i Tyskland 1933, samt honskallen från Swanscomb (200 tusen år gammal), upptäckt i England 1935. Dessa fynd tillhör bl.a. den andra interglacialen enligt det alpina schemat. Under liknande förhållanden hittades en fossil underkäke i Frankrike - Montmorin-monumentet. Dessa former kännetecknas av en liten storlek av hjärnhålan (Steingheim - 1150 cm3, Swanscombe - 1250-1300 cm3). En uppsättning egenskaper har identifierats som för de tidigaste formerna närmare den moderna människan: en relativt smal och hög skalle, en relativt konvex panna, en massiv panna, som i Pithecanthropes, inte uppdelad i beståndsdelar, en ganska rundad nacke, en uträtad panna. ansiktsregionen, närvaron av en rudimentär haka i underkäken. Det finns en tydlig arkaism i tändernas struktur: den tredje molaren är större än den andra och första (hos människor minskar storleken på molarerna från den första till den tredje). Benen hos denna art av fossil människa åtföljs av arkaiska Acheulean-verktyg.

Många kända neandertalare tillhör den sista interglaciala perioden. De tidigare levde för cirka 150 tusen år sedan. Man kan föreställa sig deras utseende från fynden från de europeiska monumenten Eringsdorf och Saccopastore. De kännetecknas av en vertikal profil av ansiktsregionen, en rundad occipital region, en försvagad superciliär relief, en ganska konvex panna, ett relativt litet antal arkaiska egenskaper i tändernas struktur (den tredje molaren är inte den största bland andra ). Hjärnvolymen hos tidiga neandertalare uppskattas till 1200-1400 cm3.
Tidpunkten för existensen av sena europeiska neandertalare sammanfaller med den senaste istiden. Den morfologiska typen av dessa former är tydligt synlig på de fossila benresterna av Chapelle (50 tusen år), Mousterian (50 tusen år), Ferrassi (50 tusen år), Neanderthal (50 tusen år), Engis (70 tusen år), Circeo (50 tusen år), San Sezer (36 tusen år) (Fig. I. 10).
Denna variant kännetecknas av en stark utveckling av ögonbrynet, den occipitala regionen komprimerad från topp till botten ("chignon-formad"), en bred näsöppning och en förstorad hålighet av molarer. Morfologer noterar närvaron av en occipital ås, ett hakutsprång (sällan och i sin linda), en stor volym av hjärnhålan: från 1350 till 1700 cm3. Enligt benen i kroppens skelett kan det bedömas att de sena neandertalarna kännetecknades av en stark, massiv kroppsbyggnad (kroppslängd - 155-165 cm). De nedre extremiteterna är kortare än hos moderna människor, lårbenen är böjda. Den breda ansiktsdelen av skallen hos neandertalarna sticker starkt fram och avfasade på sidorna, de zygomatiska benen är strömlinjeformade. Lederna i armar och ben är stora. När det gäller kroppsproportioner liknade neandertalarna den moderna eskimåtypen, vilket hjälpte dem att hålla kroppstemperaturen i kalla klimat.
Ett intressant försök görs att överföra ekologisk kunskap om den moderna människan till paleoantropologiska rekonstruktioner. Således förklaras ett antal strukturella drag hos de "klassiska" neandertalarna i Västeuropa av konsekvensen av anpassning till kalla klimatförhållanden.
Det verkar som om de tidigaste och efterföljande formerna från Europa är sammanlänkade med genetiska länkar. Europeiska neandertalare har upptäckts i Frankrike, Italien, Jugoslavien, Tyskland, Tjeckoslovakien, Ungern, Krim och norra Kaukasus.
För att lösa frågan om den moderna människans ursprung är fynden av paleoantroper utanför Europa, främst i Sydostasien och Afrika, av exceptionellt intresse. Frånvaron av egenskaper för specialisering i morfologi skiljer dem i de flesta fall från europeiska former. Så de kännetecknas av rakare och tunnare lemmar, inte så kraftfulla supraorbitala åsar, förkortade och mindre massiva skallar.
Enligt en synvinkel existerade en typisk neandertalman endast inom Europa och vissa regioner i Asien, dit han kunde flytta från Europa. Dessutom, från och med årsskiftet 40 tusen år, samexisterade neandertalarna med väletablerade människor av den moderna anatomiska typen; i Mellanöstern kan en sådan samexistens vara längre.
Fynden av paleoantroper från berget Karmel (Israel) är exceptionella i sin betydelse. De lockade forskare med en mosaik av sapiens och neandertaloiddrag. Dessa fynd kan tolkas som faktiska bevis på att tidiga neandertalare och moderna människor har blandat ihop sig. Visserligen bör det noteras att vissa Skhulfynd för närvarande anses tillhöra "arkaiska Homo sapiens". Låt oss nämna några av de mest kända fynden.
Tabun är en fossil dödskalle upptäckt i Tabun Cave, Mount Carmel. Antiken - 100 tusen år. Skallen är låg, pannan lutar, det finns supraorbitala åsar, men den främre delen och occipitalregionen har en modern karaktär. Lemmarnas böjda ben påminner om typen av europeiska neandertalare.
Skhul-V, antiken - 90 tusen år (Fig. I. 11). Skallen kombinerar en stor volym av hjärnhålan och en ganska hög panna med en modern struktur av ansiktsregionen och bakhuvudet.
Amud, antiken - 50 tusen år. Hittas i Amud-grottan nära Tiberiasjön. (Israel). Har en stor hjärnvolym: 1740 cm3. Lemmarnas ben är långsträckta.
Kafzeh, antiken - cirka 100 år. år. Öppnade i Israel. Sapience är ganska uttalad, därför anses det vara en fulländad sapiens.
I norra Irak upptäcktes en Shanidar-neandertalare, klassisk till sin typ, med en stor hjärnregion, forskarna uppmärksammade frånvaron av en kontinuerlig supraorbital ås. Ålder - 70-80 tusen år.
En upptäckt av en neandertalman med spår av en begravningsrit gjordes på Uzbekistans territorium. Skallen tillhörde en pojke med en oformad supraorbital ås. Ansiktsregionen och skelettets lemmar är enligt vissa antropologer av modern typ. Fyndplatsen är Teshchik-Tash-grottan, antiken är 70 tusen år.

På Krim, i Kiik-Koba-grottan, hittades benrester av en vuxen paleoantrop (typen är nära västeuropeiska neandertalare) och ett mycket ungt neandertalbarn. Benrester av flera neandertalbarn upptäcktes på Krim och nära staden Belogorsk. Ett fragment av skallen på en neandertalkvinna med några moderna funktioner vilket gör det likt skhulfynden. Neandertalben och tänder har upptäckts i Adygea och Georgia.
Skallen av en paleoantrop upptäcktes i Asien - i Kina, i Mala-grottan. Man tror att den inte kan hänföras till någon europeisk variant av neandertalarna. Vikten av detta fynd ligger i det faktum att det bevisar ersättningen av en scentyp med en annan i den asiatiska delen av världen. En annan synpunkt är att vi i fynd som Mala, Chanyang, Ordos (Mongolien) ser övergångsformer från Pithecanthropes till "tidiga" sapiens. Dessutom kan denna övergång i vissa former dateras till minst 0,2 miljoner år (uranmetoden).
På ungefär. Java, nära byn Ngan-dong, hittade en sorts dödskalle som bar spår av kannibalism. Forskarna uppmärksammade deras mycket tjocka väggar och kraftfulla supraorbitala ås. Sådana egenskaper gör att Ngandong-skallarna liknar Pithecanthropus-typen. Tidpunkten för existensen av de upptäckta hominiderna är övre pleistocen (cirka 0,1 miljoner år), det vill säga de är synkrona med de sena Pithecanthropes. Inom vetenskapen fanns det en åsikt att detta är en lokal, säregen typ av neandertalare, bildad som ett resultat av en långsam evolutionär process. Med andra termer definieras "Javantroperna" i Ngandong som sena Pithecanthropes genetiskt relaterade till sena Pleistocene sapiens i Australien.
Tills nyligen trodde man att neandertalarna inte bara fanns i norr, utan även i södra Afrika. Broken Hill och Saldanha hominider citerades som exempel på "syd" afrikaner. I sin morfologiska typ fann de gemensamma drag Neandertalare och Pithecanthropes. Deras hjärnvolym nådde cirka 1300 cm3 (något mindre än medelvärdet för neandertalare). Det har föreslagits att Broken Hill Man är efterträdaren till den östafrikanska Olduvai Pithecanthropus. Vissa antropologer trodde att det fanns en parallell utvecklingslinje för paleoantroper i Sydostasien och södra Afrika. För närvarande tilldelas Broken Hill-varianten rollen som en fossil sapiensform.
En förändring i taxonomiska synsätt på sena hominider har lett till det faktum att många former som föregår den moderna människan tillskrivs arkaiska Homo sapiens, som ofta förstår denna term som "pro-neandertalare" (Swanscomb, Steinheim), vidare - säregna afrikanska former (Brocken Hill , Saldanha), asiatiska (Ngandong), såväl som europeiska varianter av Pithecanthropus.
Paleontologiska bevis tyder på ett mestisursprung för klassiska europeiska neandertalare. Tydligen fanns det två vågor av migranter från Afrika och Asien för cirka 300-250 tusen år sedan, med efterföljande blandning.
Neandertalarnas evolutionära öde är inte klart. Valet av hypoteser är ganska brett: fullständig omvandling av neandertalare till sapiens; fullständig utrotning av neandertalare av sapiens av icke-europeiskt ursprung; blandning av båda alternativen. Den sista synpunkten, enligt vilken den framväxande mannen av den moderna typen migrerade från Afrika till Europa genom Asien, har det största stödet. I Asien spelades den in för cirka 100 tusen år sedan, och den kom till Europa vid 40 tusen årsskiftet. Vidare ägde assimileringen av neandertalbefolkningen rum. Bevis tillhandahålls av europeiska fynd av neandertalhominider, modern typ och mellanformer. Tidiga neandertalare, som tränger in i Mindre Asien, kunde korsas med forntida sapiens där också.
En uppfattning om omfattningen av metiseringsprocesser tillhandahålls av fossila odontologiska material. De registrerade europeiska neandertalares bidrag till den moderna människans genpool. Neandertalversionen av fossila hominider samexisterade med den moderna i tiotusentals år.
Kärnan i den evolutionära övergången som ägde rum vid gränsen till den övre paleolitikum förklaras i professor Ya.Yas hypotes. Roginsky.
Författaren sammanfattar uppgifterna om strukturen av endokranen med kliniska observationer av den moderna människan och föreslår på denna grund att paleoantropernas och den moderna människans sociala beteende skiljer sig avsevärt (beteendekontroll, manifestation av aggressivitet).
Den Mousterianska eran, som sammanfaller i tiden med eran av neandertalarnas existens, tillhör Mellanpaleolitikum. I absoluta tal given tidär från 40 till 200 tusen år. Mousteriska verktygskomplex är heterogena när det gäller förhållandet mellan verktyg annan typ. Mousteriska monument är kända i tre delar av världen - Europa, Afrika och Asien, och där upptäcktes även benresterna av neandertalarna.
Tekniken för stenbearbetning av neandertalmannen kännetecknas av en relativt hög nivå av klyvning och sekundär bearbetning av flingor. Teknikens höjdpunkt är metoden för att förbereda ytan på stenämnet och bearbeta plattorna som är separerade från den.

Noggrann korrigering av arbetsstyckets yta innebar plattornas tunnhet och perfektion av verktygen som erhölls från dem (fig. 1.12).
Den Mousterian kulturen kännetecknas av skivformade ämnen, från vilka flingorna skars av radiellt: från kanterna till mitten. Mest av Mousteriska verktyg tillverkades på flingor genom sekundär bearbetning. Arkeologer räknar dussintals typer av verktyg, men deras mångfald kommer tydligen ner på tre typer: spetsig, sidskrapa och kniv. Spetsen var ett verktyg med en spets i änden, som användes för att skära kött, läder, träbearbetning och även som dolk eller spjutspets. Skrapan var en flinga, retuscherad längs kanten. Detta verktyg användes för att skrapa eller skära vid bearbetning av slaktkroppar, skinn eller trä. Trähandtag lades till skraporna. Tandade verktyg användes för att svarva träföremål, för att skära eller såga. Det finns piercer, framtänder, skrapor i Mousterian - verktyg från den sena paleolitikum. Arbetsmedel representeras av speciella flishuggare (bitar av sten eller småsten med en långsträckt form) och retuschörer (bitar av sten eller ben för bearbetning av verktygets kant genom att trycka).
Moderna etnografiska studier av australiensiska aboriginerna hjälper till att presentera stenålderns tekniska processer. Arkeologernas experiment har visat att tekniken för att få verktygsämnen i form av flingor och plattor var komplex och krävde erfarenhet, teknisk kunskap, exakt koordination av rörelser och stor uppmärksamhet.
Erfarenhet gjorde det möjligt för den forntida människan att minska den tid som behövdes för att tillverka verktyg. Benbearbetningstekniken i Mousterian är dåligt utvecklad. Träverktyg användes i stor utsträckning: klubbor, spjut, horn med ändar härdade i eld. Kärl för vatten och delar av bostäder var gjorda av trä.
Neandertalarna var skickliga jägare. På deras platser hittades ansamlingar av ben från stora djur: mammutar, grottbjörnar, bison, vilda hästar, antiloper, bergsgetter. Komplexa jaktaktiviteter låg inom kraften av ett samordnat team av neandertalare. Mousterianerna använde metoder för att runda upp eller få spår av djur till raster och träsk. Sammansatta verktyg hittades - spjutspetsar med flintfragment. Bolas användes som kastvapen. Mousterianerna övade på att skära upp slaktkropparna av slaktade djur och steka köttet över eld. De gjorde enkla kläder åt sig själva. Att samlas var av viss betydelse. De upptäckta rivjärnen i sten antyder att det förekom en primitiv bearbetning av spannmål. Kannibalism fanns bland neandertalare, men var inte utbredd.
Under Mousterian-tiden förändrades bosättningarnas karaktär. Bodar, grottor och grottor var oftare bebodda. Typer av neandertalbosättningar urskiljs: verkstäder, jakt och basläger. För att skydda bränderna från vinden anordnades vindskydd. I grottorna gjordes trottoarer av småsten och bitar av kalksten.
Benrester av neandertalare kan hittas tillsammans med verktyg från övre paleolitikum, vilket till exempel var fallet med upptäckten av en sen paleoantrop i Frankrike (Saint-Cezaire-platsen).
Under den tidiga Wurms tidevarv dök Mousterian-begravningar upp på Eurasiens territorium - de första pålitliga spåren av de dödas begravning. Idag har ett 60-tal sådana monument upptäckts. Intressant nog begravde grupperna "Neanderthaloid" och "sapient" oftare vuxna, medan "neandertalarna" begravde både vuxna och barn i samma utsträckning. Fakta om begravningen av de döda ger anledning att anta existensen av en dualistisk världsbild bland Mousterianerna.
Modern människa, fossil och modern (neoantroper)
Fossila representanter för Homo sapiens sapiens är brett representerade i de kända arkeologiska fynden av hominidlämningar. Den maximala geologiska åldern för neoantroper som är helt bildade i utvecklingen av fossiler uppskattades tidigare till cirka 40 tusen år (ett fynd i Indonesien). Man tror nu att de sapiens som hittats i Afrika och Asien var av mycket äldre forntid (även om vi talar om skelett som har arkaiska drag uttryckta i varierande grad).
Benresterna av en fossil man av denna underart är utbredd: från Kalimantan till Europas yttersta delar.
Namnet "Cro-Magnon" (som fossila neoantroper hänvisas till i litteraturen) beror på det berömda franska monumentet av den övre paleolitiska Cro-Magnon. Strukturen av skallen och skelettet i kroppen av fossila neoantroper skiljer sig inte i princip från moderna människor, även om dess ben är mer massiva.
Enligt analysen av benmaterial från senpaleolitiska begravningar, genomsnittlig ålder Cro-Magnons var 30-50 år gammal. Samma förväntade livslängd bevarades fram till medeltiden. Patologi hos ben och tänder är mindre vanligt än skador (Cro-Magnon-tänderna var friska).
Tecken på skillnad mellan skallen hos Cro-Magnons och Neanderthals (Fig. 1.13): en mindre utstående ansiktsregion, en hög konvex krona, en hög rak panna, en rundad nackknöl, mindre fyrkantiga ögonhålor, en mindre total storlek på skallen , ett hakutsprång av skallen bildas; den superciliära åsen är frånvarande, käkarna är mindre utvecklade, tänderna har en liten hålighet. Den största skillnaden mellan Cro-Magnons och Neandertalare är strukturen hos endokranen. Paleoneurologer tror att under sen antropogenes utvecklades de främre regionerna i hjärnan, inklusive centra för att kontrollera beteendet. Var komplicerade intern kommunikation hjärnan, men hjärnans totala storlek minskade något. Cro-Magnons var högre (169-177 cm) och mindre grovbyggda än neandertalarna.

Skillnader mellan Cro-Magnon-skallar och moderna: höjden på valvet är lägre, de längsgående dimensionerna är större, de superciliära bågarna är uttalade, ögonhålorna är bredare, ansiktsdelen av skallen och underkäken är bredare, skallens väggar är tjockare. Den övre paleolitiska mannen behöll under lång tid tecknen på det tandsystem som är karakteristiskt för neandertalaren. De egenskaper som skiljer Cro-Magnon-skallen och endokran från moderna människor är ofta "neandertaloid" i karaktär.
Uppmärksamhet dras till det faktum att Cro-Magnon-mannens distributionsområde är enormt: hela ekumenen. Med tillkomsten av Cro-Magnon-människan, enligt många experter, är arternas utveckling fullbordad, och utvecklingen av människans biologiska egenskaper i framtiden verkar omöjlig.
De mest kompletta fynden av Cro-Magnon-skelett i Europa har en antiken som inte överstiger 40 tusen år. Till exempel levde den franska neoantropen Cro-Magnon för 30 tusen år sedan, Cro-Magnon Sungir (ett distrikt i Vladimir) är 28 tusen år gammalt. Afrikas arkaiska sapiens (med ganska uttalade neandertaloiddrag) ser mycket äldre ut: Omo i Etiopien - 130 tusen år, River Mouse (Sydafrika) - 120 tusen år, Border (Sydafrika) - mer än 70 tusen år, kenyanska fynd av sapiens - 200-100 tusen år, Mumba (Tanzania) - 130 tusen år, etc. Det antas att antiken för afrikanska sapiens kan vara ännu större. Asiatiska fynd av sapiens har följande ålder: Dali (PRC) - 200 tusen år, Jinnbshan (PRC) - 200 tusen år, Qafzeh (Israel) - mer än 90 tusen år, Skhul V (Israel) - 90 tusen år, Nia ( Kalimantan) - 40 tusen år. De australiska fynden är cirka 10 tusen år gamla.
Tidigare antogs det att moderna människor uppstod i Europa för cirka 40 tusen år sedan. I dag Mer antropologer och arkeologer placerar sapiens förfäders hem i Afrika, och antiken för de senare ökar kraftigt, med fokus på ovanstående fynd. I enlighet med den tyske antropologen G. Breuers hypotes uppträdde Homo sapiens sapiens söder om Sahara för cirka 150 tusen år sedan, och migrerade sedan till Mindre Asien (på nivån 100 tusen år) och vid årsskiftet 35-40 tusen år började befolka Europa och Asien, korsning med lokala neandertalare. Moderna biomolekylära data tyder också på att den moderna mänsklighetens förfäder kom från Afrika.
I enlighet med moderna evolutionära åsikter är den mest troliga modellen "nettoutvecklingen" av hominider, där en viktig plats ges åt utbyte av gener mellan olika underarter och arter av forntida människa. Därför väldigt tidiga fynd sapiens i Afrika och Europa tolkas som bevis på korsning mellan sapiens-arter och Pithecanthropus. I processen att bli en sapiens-typ mellan de primära utvecklingscentra för släktet Homo (västra och östliga) förekom ett konstant utbyte av gener.
För cirka 40 tusen år sedan började den snabba bosättningen av neoantropen. Orsakerna till detta fenomen ligger i människans genetik och utvecklingen av hennes kultur.
Forskare som studerar Cro-Magnon-människan måste hantera en mängd olika typer. Enhällig uppfattning om tidpunkten för tillägget moderna raser Nej. Enligt en synvinkel finns egenskaperna hos moderna raser i övre paleolitikum. Denna synvinkel illustreras av exempel på den geografiska fördelningen av två funktioner - näsans utsprång och graden av horisontell profilering av ansiktsregionen. Enligt en annan synvinkel tar raser form sent, och befolkningen i övre paleolitikum kännetecknades av stor polymorfism. Så för Europa särskiljs ibland cirka 8 typer av raser från övre paleolitikum. Två av dem ser ut så här: a) en dolichokraniell, storhövdad version av Cro-Magnon med en måttlig bredd i ansiktet och en smal näsa; b) brakykraniell (korthårig), med en mindre skalle, ett mycket brett ansikte och en bred näsa. Man kan anta att det fanns tre stadier i bildandet av raser: 1) Mellan- och Nedre paleolitikum - bildandet av vissa rasegenskaper; 2) Övre paleolitikum - början på bildandet av raskomplex; 3) post-paleolitisk tid - tillägg av raser.
Kulturerna i den övre (sena) paleolitikum är förknippade med utseendet på moderna människor (neoantroper). I Europa förra perioden Paleolitikum (gammal stenålder) beräknas vara 35-10 tusen år före våra dagar och sammanfaller med tiden för den sista Pleistocene glaciationen (detta faktum är föremål för diskussion i samband med problemet med miljöns roll i utvecklingen av mänskligheten) (Fig. I. 14).
Vid första anblicken, under den paleolitiska eran som diskuteras, fanns det inga grundläggande skillnader i materiell kultur från tidigare epoker: samma stenredskap och jaktredskap. Faktum är att Cro-Magnons tillverkade en mer komplex uppsättning verktyg: knivar (ibland dolkar), spjutspetsar, mejselskärare, benverktyg som sylar, nålar, harpuner etc. Benverktyg stod för ungefär hälften av hela lagret, de var starka och mer hållbara än sten. Stenverktyg användes för att göra verktyg av ben, trä, elfenben - så här var de tekniska kedjorna i den antika människans handlingar komplicerade.

Helt nya typer av redskap uppstod, som nålar med ögon, fiskkrokar, harpuner, spjutkastare. De ökade avsevärt människans makt över naturen.
Den största skillnaden mellan den övre paleolitikum var förbättringen av stenbearbetningen. Under Moustertiden fanns det flera sätt att bearbeta en tom sten (kärna). Den lavallusiska tekniken med noggrann initial ytbehandling av arbetsstycket är utgångspunkten för tekniken från den övre paleolitikum. Cro-Magnons använde ämnen som lämpade sig för flisning av en serie plattor (prismatiska kärnor). Sålunda, under den övre paleolitiska eran, förbättrades flisningsteknikerna, vilket resulterade i högkvalitativa mikroblad lämpliga för användning i kompositverktyg.
Arkeologer har experimenterat med att rekonstruera hur plattorna är separerade från kärnan, som Cro-Magnons gjorde. Den utvalda och speciellt bearbetade kärnan klämdes fast mellan knäna, som spelade rollen som en stötdämpare. Separationen av plattorna utfördes med användning av en stenhuggare och en benmellanprodukt. Dessutom separerades flintplattor genom att man tryckte på kärnans kant med en ben- eller stenvridare.
Knivbladsmetoden är mycket mer ekonomisk än flingmetoden. Från ett arbetsstycke kunde en skicklig hantverkare separera mer än 50 plattor på kort tid (längd upp till 25-30 cm och tjocklek - flera millimeter). Arbetseggen på det knivliknande bladet är mycket större än flingans. Mer än 100 typer av verktyg är kända för senpaleolitikum. Det föreslås att olika Cro-Magnon-verkstäder kan skilja sig åt i originaliteten hos tekniskt "mode".
I övre paleolitikum var jakten ännu mer perfekt än under Moustertiden. Detta spelade en stor roll för att öka matresurserna och i samband med detta befolkningen.
En perfekt innovation var spjutkastare, som gav Cro-Magnon-handen en styrka, vilket fördubblade avståndet över vilket ett spjut kunde kastas (upp till 137 m, med ett optimalt avstånd för att slå upp till 28 m). Harpuner gjorde det möjligt att fånga fisk effektivt. Cro-Magnon uppfann snaror för fåglar, fällor för djur.
Perfekt jakt utfördes på ett stort vilt: renar och stenbockar förföljdes under sina säsongsmässiga vandringar till nya betesmarker och tillbaka. Jakttekniker med kunskap om området - driven jakt - gjorde det möjligt att döda djur i tusental. Således bildades för första gången en oavbruten källa till mycket näringsrik mat. En person fick möjlighet att bo i svåråtkomliga områden.
Vid byggandet av bostäder använde Cro-Magnonerna Mousterians prestationer och förbättrade dem. Detta tillät dem att överleva under förhållandena under det sista kalla årtusendet av Pleistocene.
Europeiska Cro-Magnons använde sin goda kunskap om området för att bebo grottorna. Många grottor hade tillgång till söder, så de var väl uppvärmda av solen och var skyddade från kalla nordliga vindar. Grottorna valdes inte långt från vattentäkter, med god utsikt över betesmarker där hjordar av klövvilt betade. Grottorna kunde användas året runt eller för säsongsbetonade vistelser.
Cro-Magnonerna byggde också bostäder i floddalarna. De var gjorda av sten eller grävde ur marken, väggarna och taket var gjorda av skinn, och stöden och botten kunde fodras med tunga ben och betar. Den övre paleolitiska strukturen på Kostenki-orten (ryska slätten), 27 m lång, markeras av ett antal härdar i mitten, vilket indikerar att flera familjer övervintrade här.
Nomadjägare byggde lätta hyddor. svår klimatförhållanden Cro-Magnons fick hjälp att bära varma kläder. Avbildningar av människor på benartefakter tyder på att de bar åtsittande byxor för att hålla dem varma, parkas med huvor, stövlar och vantar. Klädernas sömmar var välsydda.
Cro-Magnons höga intellektuella utveckling och psykologiska komplexitet bevisas av förekomsten av många monument av primitiv konst, som är känd för en period på 35-10 tusen år i Europa. Detta syftar på små skulpturer och väggmålningar i grottor. Gravyrer av djur och människor gjordes på stenar, ben och hjorthorn. Skulpturer och basreliefer gjordes av lera och sten, och teckningar erhölls av Cro-Magnons med hjälp av ockra, mangan och träkol. Syftet med primitiv konst är inte klart. Man tror att det var av rituell karaktär.
Riklig information om Cro-Magnons liv tillhandahålls av studier av begravningar. Man fann till exempel att den förväntade livslängden för Cro-Magnon-människan ökade i jämförelse med neandertalarna.
Vissa ritualer av Cro-Magnons har rekonstruerats. Så seden att beströja den avlidnes skelett med röd ockra vittnar tydligen om tron på livet efter detta. Begravningar med rika dekorationer tyder på uppkomsten av rika människor bland jägare-samlare.
Ett utmärkt exempel på en Cro-Magnon-begravning tillhandahålls av Sungir-monumentet nära staden Vladimir. Åldern för begravningen är cirka 24-26 tusen år. Här vilar en gammal man ("Leader") i pälskläder, rikt dekorerad med pärlor. Den andra begravningen är intressant - en parad barns. Barnens skelett åtföljdes av mammutbetsspjut och var prydda med elfenbensringar och armband; kläder är också dekorerade med pärlor.
Modern människa och evolution
Sedan bildningen av Homo sapiens-arten slutfördes (från mitten av övre paleolitikum) har den behållit stabilitet i sin biologiska status. Den evolutionära fullständigheten hos en person är relativ och betyder inte ett fullständigt upphörande av förändringar i hans biologiska egenskaper. En mängd olika förändringar i den anatomiska typen av en person av den moderna typen har studerats. Exempel är en minskning av skelettets massivitet, storleken på tänderna, en förändring av småtårna etc. Man antar att dessa fenomen beror på slumpmässiga mutationer. Vissa antropologer, baserat på anatomiska observationer, förutspår utseendet av Homo futurus - "Framtidens man", med ett stort huvud, reducerat ansikte och tänder, med färre fingrar. Men dessa anatomiska "förluster" kännetecknar inte alla mänskliga populationer. En alternativ syn är att den moderna människans biologiska organisation gör det möjligt för obegränsat social evolution, så det är osannolikt att det kommer att förändras som art i framtiden.
Den evolutionära teorin om mänsklig utveckling, som den engelske vetenskapsmannen C. Darwin föreslog, blev en verklig sensation i vetenskapliga världen. Fram till dess var hela världen i full tillit till att människan är Guds skapelse. Den darwinistiska teorin, till skillnad från andra versioner av människans ursprung, kunde tydligt förklara hur hennes evolution skedde.
Ch. Darwins evolutionsteori
Mänskligheten har länge försökt reda ut mysteriet med dess utseende på planeten, men det enda svaret var bara i religionen, enligt vilken människan är en manifestation av Guds plan.
Denna förklaring passade människor tills de började aktivt utvecklas och expandera vetenskaplig kunskap. Forskare har länge kämpat för att reda ut människans ursprung, men bara den brittiske naturforskaren Charles Darwin lyckades.

Ris. 1. Charles Darwin.
Hans revolutionära teori för dessa tider, enligt vilken människan härstammade från primater, orsakade verklig dissonans i samhället. Inte alla forskare, för att inte tala om vanliga människor, ville se apor bland sina gamla förfäder.
Darwins teori presenterade dock en hel del tungt vägande bevis. För mycket kopplade människan till djurvärlden: skelettets struktur, nervsystem, matsmältnings-, cirkulations- och andningsorgan. Den största likheten hos människor var med primater.
TOP 4 artiklarsom läser med detta
Den viktigaste förutsättningen för "humanisering" av primater var användningen av naturliga föremål som ett verktyg för skydd mot fiender eller jakt på vilda djur.

Ris. 2. Primitiva verktyg.
De viktigaste stadierna av mänsklig evolution
Bearbeta evolutionär utveckling mänskligheten, från primater till modern människa, tog flera miljoner år. Totalt finns det fem huvudstadier av mänsklig evolution, som var och en har sina egna särdrag.
I hjärtat av alla evolutionära processer finns den viktigaste naturlagen - det naturliga urvalet, tack vare vilket arten får möjlighet att anpassa sig så bra som möjligt till miljöförhållandena.

Ris. 3. Primitivt samhälle.
Tabell "Stadier av mänsklig evolution"
|
Stadier av mänsklig evolution |
Strukturella egenskaper |
Livsstil |
Verktyg |
|
Stora apor - Australopithecus |
Höjd 120-140 cm Skallevolym - 500-600 kubikmeter. cm, upprätt hållning |
De byggde inte permanenta bostäder, använde inte eld, livsstilen är flock |
Pinnar och stenar |
|
Forntida människor - smart man |
Hjärnans volym är 680 kubikmeter. centimeter, |
Visste inte hur man använder eld |
Verktyg i form av stenar med spetsiga kanter |
|
De äldsta människorna - Homo erectus (Pithecanthropus, Sinanthropus, Heidelbergman) |
Höjd 170 cm.Hjärnvolym - 900-110 kubikmeter. se Foten har en båge, höger hand är bättre utvecklad, konstant upprätt hållning, förändringar i käkapparaten, utseendet av kurvor i ryggraden |
De stödde elden, byggde bostäder, jagade tillsammans. Det fanns rudiment av artikulerat tal |
Olika verktyg gjorda av sten, bland vilka det viktigaste är en stenyxa |
|
Forntida människor - Neandertalare |
Höjd 156 cm.Hjärnvolym - 1400 kubikmeter. se Det finns en rudiment av hakans utsprång, en utvecklad hand, en välvd fot, en hög skallbåge, en inte så massiv underkäke |
De kunde bygga bostäder, producera och underhålla eld. Boende i grupper om 50-100 personer. |
Arbetsredskap är mycket olika: sidskrapor, spetsar gjorda av sten, ben och trä |
|
De första moderna människorna - Cro-Magnons |
Höjd 180 cm, hjärnvolym - 1600 kubikmeter. se Utseende som är inneboende i den moderna människan |
Utvecklat tal, religionens och konstens början, förmågan att göra kläder dök upp. Bor i bosättningar som en del av ett stamsamhälle. Utveckling av jordbruk och djurhållning |
För tillverkning av verktyg används ett brett utbud av material: trä, ben, stenar, horn. Spjut, pilar, knivar, skrapor gjordes av dem. |
Vad har vi lärt oss?
När vi studerade ämnet "Tabell" Stadier av mänsklig evolution "i 11:e klassprogrammet lärde vi oss vilken teori som indikerar människans ursprung från primater och vilka evolutionsstadier en person måste gå igenom för att nå toppen av sin utveckling .
Ämnesquiz
Rapportutvärdering
Genomsnittligt betyg: 4.3. Totalt antal mottagna betyg: 235.
Antropogenes (grek. anthropos man, génesis ursprung), del biologisk evolution, vilket ledde till uppkomsten av arten Homo sapiens, som separerade från andra hominider, antropoid
apor och placenta däggdjur. Detta är processen för historisk och evolutionär bildning av den fysiska typen av en person, den initiala utvecklingen av hans arbetsaktivitet, tal och samhälle.
Stadier av mänsklig evolution
Forskare hävdar att den moderna människan inte härstammar från moderna antropoida apor, som kännetecknas av en snäv specialisering (anpassning till en strikt definierad livsstil i tropiska skogar), utan från högorganiserade djur som dog ut för flera miljoner år sedan - driopithecus.
Enligt paleontologiska fynd (fossiler), för cirka 30 miljoner år sedan, uppträdde forntida parapithecus-primater på jorden, som levde i öppna ytor och på träd. Deras käkar och tänder liknade människoapor. Parapithecus gav upphov till moderna gibboner och orangutanger, samt en utdöd gren av driopithecus. De senare i sin utveckling var uppdelade i tre linjer: en av dem ledde till den moderna gorillan, den andra till schimpansen och den tredje till Australopithecus och från den till människan. Förhållandet mellan driopithecus och människan etablerades på grundval av en studie av strukturen hos hans käke och tänder, upptäckt 1856 i Frankrike. Det viktigaste steget i omvandlingen av apliknande djur till de äldsta människorna var utseendet på tvåfotingsrörelse. I samband med klimatförändringar och gallring av skog har det skett en övergång från ett trädlevande till ett terrestriskt sätt att leva; för att bättre kunna se området där människans förfäder hade många fiender, var de tvungna att stå på bakbenen. Därefter utvecklade det naturliga urvalet och fixerade upprätt hållning, och som ett resultat av detta befriades händerna från funktionerna stöd och rörelse. Så här uppstod australopithecines - släktet som hominider tillhör (en familj av människor).
australopitheciner
Australopithecus - högt utvecklade tvåfota primater som använde naturliga föremål som verktyg (därav kan australopitheciner ännu inte betraktas som människor). Benrester av Australopithecus upptäcktes först 1924 i Sydafrika. De var lika stora som en schimpans och vägde cirka 50 kg, hjärnvolymen nådde 500 cm3 - på grundval av detta är Australopithecus närmare människor än någon av de fossila och moderna aporna.
Strukturen på bäckenbenen och huvudets position liknade en persons, vilket indikerar en uträtad position av kroppen. De levde för cirka 9 miljoner år sedan i öppna stäpper och livnärde sig på växt- och djurfoder. Verktygen för deras arbete var stenar, ben, pinnar, käkar utan spår av konstgjord bearbetning.
skicklig man
Utan att ha en snäv specialisering av den allmänna strukturen, gav Australopithecus upphov till en mer progressiv form, kallad Homo habilis - en skicklig man. Dess benrester upptäcktes 1959 i Tanzania. Deras ålder bestäms till cirka 2 miljoner år. Tillväxten av denna varelse nådde 150 cm. Hjärnans volym var 100 cm3 större än den hos Australopithecus, tänderna hos en mänsklig typ, fingrarnas falanger, som hos en person, är tillplattade.
Även om det kombinerade tecken på både apor och människor, indikerar övergången av denna varelse till tillverkning av stenverktyg (välgjorda stenar) utseendet på arbetsaktivitet i den. De kunde fånga djur, kasta sten och utföra andra aktiviteter. De benhögar som hittats tillsammans med fossilerna av Homo sapiens vittnar om det faktum att kött har blivit en permanent del av deras kost. Dessa hominider använde grova stenredskap.
Homo erectus
Homo erectus - Homo erectus. arten som den moderna människan tros härstamma från. Dess ålder är 1,5 miljoner år. Hans käkar, tänder och ögonbrynsryggar var fortfarande massiva, men hjärnvolymen hos vissa individer var densamma som den moderna människan.
Några ben av Homo erectus har hittats i grottor, vilket tyder på ett permanent hem. Förutom djurben och ganska välgjorda stenredskap hittades högar av träkol och brända ben i några grottor, så att Australopithecus tydligen redan vid denna tid hade lärt sig hur man gör upp eld.
Detta stadium av homininutveckling sammanfaller med koloniseringen av andra kallare regioner av afrikaner. Det skulle vara omöjligt att överleva de kalla vintrarna utan att utveckla komplexa beteenden eller tekniska färdigheter. Forskare antyder att den förmänskliga hjärnan hos Homo erectus kunde hitta sociala och tekniska lösningar (eld, kläder, matförsörjning och samlevnad i grottor) på problemen i samband med behovet av att överleva i vinterkylan.
Således anses alla fossila hominider, särskilt Australopithecus, vara föregångare till människor.
Evolution fysiska egenskaper de första människorna, inklusive den moderna människan, täcker tre stadier: forntida människor, eller arkantroper;forntida människor, eller paleoantroper;moderna människor eller neoantroper.
arkantroper
Den första representanten för arkantroperna är Pithecanthropus (japansk man) - en apmänniska, upprätt. Hans ben hittades ca. Java (Indonesien) 1891. Ursprungligen bestämdes dess ålder till 1 miljon år, men enligt en mer exakt modern uppskattning är den lite över 400 tusen år gammal. Pithecanthropus höjd var cirka 170 cm, kraniets volym var 900 cm3. Något senare fanns det en synantrop (kinesisk man). Många rester av den hittades under perioden 1927 till 1963. i en grotta nära Peking. Denna varelse använde eld och gjorde stenverktyg. Till denna grupp forntida människor hör också Heidelbergmannen.
Paleoantroper
Paleoantroper - Neandertalarna verkade ersätta arkantroperna. För 250-100 tusen år sedan var de allmänt bosatta i Europa. Afrika. Front och Sydasien. Neandertalarna gjorde en mängd olika stenredskap: handyxor, sidoskrapor, spetsiga; begagnad eld, grova kläder. Volymen på deras hjärna växte med 1400 cm3.
Funktioner i strukturen i underkäken visar att de hade rudimentärt tal. De levde i grupper om 50-100 individer och under glaciärernas uppkomst använde de grottor och drev ut vilda djur ur dem.
Neoantroper och Homo sapiens
Neandertalarna ersattes av människor av den moderna typen - Cro-Magnons - eller neoantroper. De dök upp för cirka 50 tusen år sedan (deras benrester hittades 1868 i Frankrike). Cro-Magnons utgör det enda släktet och arten av Homo Sapiens - Homo sapiens. Deras apdrag var helt utjämnade, det fanns ett karakteristiskt hakutsprång på underkäken, vilket tyder på deras förmåga att artikulera tal, och i konsten att göra olika verktyg av sten, ben och horn, hade Cro-Magnons gått långt före jämfört med till neandertalarna.
De tämjde djur och började bemästra jordbruket, vilket gjorde det möjligt att bli av med hunger och få i sig en mängd olika livsmedel. Till skillnad från deras föregångare skedde utvecklingen av Cro-Magnon-folket under stort inflytande av sociala faktorer (teambuilding, ömsesidigt stöd, förbättring av arbetsaktiviteten, en högre nivå av tänkande).
Uppkomsten av Cro-Magnons är det sista steget i bildandet av en modern typ av person . Den primitiva mänskliga flocken ersattes av det första stamsystemet, som fullbordade bildandet av det mänskliga samhället, vars fortsatta framsteg började bestämmas av socioekonomiska lagar.
18) Bevis för människans ursprung från djur. Atavismer och rudiment hos människan.
TILL de hänvisas traditionellt till jämförande anatomiska, embryologiska, fysiologiska och biokemiska, molekylärgenetiska, paleontologiska.
1. Jämförande anatomisk.
Den allmänna planen för människokroppens struktur liknar strukturen av kordatkroppen. Skelettet består av samma sektioner som hos andra däggdjur. Kroppshålan delas av diafragman i buk- och bröstregionerna. Nervsystemet av tubulär typ. I mellanörat finns tre hörselben (hammare, städ, stigbygel), det finns öron och öronmuskler förknippade med dem. I mänsklig hud, liksom andra däggdjur, finns mjölk-, talg- och svettkörtlar. Cirkulationssystemet är stängt, det finns ett fyrkammarhjärta. Bekräftelse på människans animaliska ursprung är närvaron av rudiment och atavismer i henne.
2. Embryologisk.
I mänsklig embryogenes observeras de viktigaste utvecklingsstadierna som är karakteristiska för ryggradsdjur (krossning, blastula, gastrula etc.) I de tidiga stadierna av embryonal utveckling har det mänskliga embryot tecken som är karakteristiska för lägre ryggradsdjur: korda, gälskåror i svalget kavitet, ihåligt nervrör, bilateral symmetri i kroppens struktur, slät yta av hjärnan. Ytterligare utveckling av embryot visar egenskaper som är karakteristiska för däggdjur: flera par bröstvårtor, närvaron av hår på kroppsytan, som hos alla däggdjur (utom monotremes och pungdjur), utvecklingen av ungen inuti moderns kropp och näring av fostret genom moderkakan.
3. Fysiologiska och biokemiska.
Hos människor och människoapor är strukturen av hemoglobin och andra kroppsproteiner mycket nära. Det finns en likhet i blodgrupper. Blodet från pygméschimpansen (bonobo) i motsvarande grupp kan också transfunderas till människor. Det finns också ett blod Rh-antigen hos människor (det identifierades först i Rhesus-apan). Människor är nära människor under graviditeten, tidpunkten för puberteten.
4. Molekylärgenetisk.
Alla människoapor har ett diploid antal kromosomer 2 n = 48. Hos människor är 2 n = 46 (det har fastställts att mänsklig kromosom 2 bildas genom sammansmältning av två kromosomer homologa med de hos schimpanser). Tillgängligt hög grad homologi av primär struktur gener (mer än 90 % av mänskliga och schimpansgener liknar varandra).
5. Paleontologisk.
Många fossila lämningar har hittats (individuella ben, tänder, fragment av skelettet, verktyg etc.), som gör det möjligt att komponera en evolutionär serie av förfäders former av den moderna människan och förklara de viktigaste riktningarna för deras utveckling.
Skillnaden mellan människa och djur
De ärftliga förändringar som uppstod under evolutionens gång under kontroll av naturligt urval bidrog till uppkomsten av en upprätt hållning hos människor, frigörande av händer, utveckling och ökning av hjärnskallen och minskningen av dess ansiktsdel. Samtidigt utvecklade en person ett behov av systematisk tillverkning av verktyg, vilket bidrog till att förbättra strukturen och funktionen hos handen, hjärnan, talapparaten, mental aktivitet och uppkomsten av tal. En betydande roll i utvecklingen av hjärnan och handen spelades av binokulär (stereoskopisk) färgseende, som fanns hos mänskliga förfäder.
Atavismer och rudiment hos människan.
Rudiment är organ som har förlorat sin huvudsakliga betydelse i processen för den evolutionära utvecklingen av organismen.
Många rudimentella organ är inte helt värdelösa och utför någon sekundär funktion med hjälp av strukturer som tydligen är avsedda för mer komplexa syften.
Atavism är utseendet hos en individ av tecken som är karakteristiska för avlägsna förfäder, men frånvarande i de närmaste.
Uppkomsten av atavismer förklaras av det faktum att generna som är ansvariga för denna egenskap bevaras i DNA, men fungerar inte, eftersom de undertrycks av andra geners verkan.
Rudiment hos människor:
∙ svanskotor;
∙ vissa människor har en rudimentiell stjärtmuskel, extensor coccygis, som är identisk med de muskler som rör svansen hos andra däggdjur. Den är fäst vid svanskotan, men eftersom svanskotan hos människor praktiskt taget inte kan röra sig är denna muskel värdelös för människor;
∙ kroppshårfäste;
∙ speciella muskler arrectores pilorum, som i våra förfäder tjänade till att "höja håret på slutet" (detta är användbart för termoreglering, och hjälper också djur att se större ut - för att skrämma rovdjur och konkurrenter). Hos människor leder sammandragningen av dessa muskler till "gåshud", vilket knappast kan vara det något adaptivt värde;
∙ tre öronmuskler som gjorde att våra förfäder kunde röra sina öron. Det finns människor som vet hur man använder dessa muskler. Detta hjälper djur med stora auriklar att bestämma riktningen för ljudkällan, men hos människor kan denna förmåga bara användas för skojs skull;
∙ blinkers ventriklar i struphuvudet;
∙ blindtarmens blindtarm (appendix). Långtidsobservationer har visat att borttagandet av blindtarmen inte har någon betydande inverkan på människors förväntade livslängd och hälsa, förutom det faktum att människor efter denna operation i genomsnitt blir sjuka av kolit lite mindre ofta;
∙ en gripreflex hos nyfödda (det hjälper apungar att hålla fast vid sin mammas päls);
∙ hicka: vi ärvde denna reflexrörelse från våra avlägsna förfäder - amfibier. I en grodyngel låter denna reflex dig snabbt passera en del vatten genom gälskårorna. Hos både människor och grodyngel styrs denna reflex av samma del av hjärnan och kan undertryckas på samma sätt (till exempel inandning koldioxid eller expansion av bröstet)
∙ lanugo: ett hårfäste som utvecklas i det mänskliga embryot på nästan hela kroppen, förutom handflatorna och fötterna, och försvinner strax före födseln (för tidigt födda barn föds ibland med lanugo).
Exempel på atavismer:
∙ kaudalt bihang hos människor;
∙ kontinuerlig hårfäste på människokroppen;
∙ ytterligare par bröstkörtlar;
nitton . Åldrande av kroppen. Teorier om åldrande. Geriatrik och gerontologi.
Ålderdom är ett stadium av individuell utveckling, när det når vilket regelbundna förändringar observeras i kroppen i det fysiska tillståndet, utseendet, den känslomässiga sfären Senila förändringar blir uppenbara och ökar under den post-reproduktiva perioden av ontogenes. Början av nedgången av den reproduktiva funktionen eller till och med dess fullständiga förlust kan dock inte tjäna som den nedre gränsen för ålderdom. Faktum är att klimakteriet hos kvinnor, som består i att upphöra med frisättningen av mogna ägg från äggstocken och följaktligen upphörandet av menstruationsblödning, bestämmer slutet på den reproduktiva perioden av livet. Samtidigt, när klimakteriet nås, är de flesta funktioner och yttre tecken långt ifrån att nå det tillstånd som är karakteristiskt för gamla människor. Å andra sidan börjar många av de förändringar vi förknippar med ålderdom före reproduktiv nedgång. Detta gäller både fysiska tecken (grånande hår, utveckling av framsynthet) och olika organs funktioner. Till exempel, hos män, börjar en minskning av frisättningen av manliga könshormoner från gonaderna och en ökning av frisättningen av gonadotropa hormoner från hypofysen, vilket är typiskt för en gammal organism, vid cirka 25 års ålder.
Det finns kronologisk och biologisk (fysiologisk) ålder.
Enligt den moderna klassificeringen, baserat på bedömningen av många genomsnittliga indikatorer på kroppens tillstånd, kallas personer vars kronologiska ålder har nått 60-74 år gamla, 75-89 år gamla, över 90 år gamla - hundraåringar. En exakt bestämning av biologisk ålder är svår eftersom individuella ålderstecken uppträder i olika kronologiska åldrar och kännetecknas av olika tillväxthastigheter. Dessutom är åldersrelaterade förändringar i ens ett tecken föremål för betydande sexuella och individuella fluktuationer.
Låt oss överväga ett sådant tecken som elasticitet (elasticitet) i huden. Samma biologiska ålder uppnås i det här fallet av en kvinna vid cirka 30 år och av en man vid 80. Det är därför, först och främst, kvinnor behöver kompetent och konstant hudvård. För att bestämma den biologiska åldern, som är nödvändig för att bedöma åldringshastigheten, används testbatterier som gör en kumulativ bedömning av många tecken samtidigt som naturligt förändras under livets gång.
Sådana batterier är baserade på komplexa funktionella indikatorer, vars tillstånd beror på den samordnade aktiviteten hos flera kroppssystem. Enkla tester är vanligtvis mindre informativa. Till exempel minskar hastigheten för utbredning av en nervimpuls, som beror på nervfiberns tillstånd, med 10 % i åldersintervallet 20-90 år, medan vital kapacitet lungor, bestäms av det samordnade arbetet i andnings-, nerv- och muskelsystemen, med 50%.
Ålderdomstillståndet uppnås genom de förändringar som utgör innehållet i åldringsprocessen. Denna process fångar alla nivåer av en individs strukturella organisation - molekylär, subcellulär, cellulär, vävnad, organ. Det kumulativa resultatet av många speciella manifestationer av åldrande på nivån för hela organismen är en ökande minskning av en individs livsduglighet med åldern, en minskning av effektiviteten hos adaptiva, homeostatiska mekanismer. Det har till exempel visat sig att unga råttor efter nedsänkning i isvatten i 3 minuter återställer sin kroppstemperatur på cirka 1 h. Medelålders djur behöver 1,5 timmar för detta och cirka 2 timmar för gamla.
I allmänhet leder åldrande till en progressiv ökning av sannolikheten för dödsfall. Den biologiska innebörden av åldrande är alltså att det gör organismens död oundviklig. Det senare är ett universellt sätt att begränsa deltagandet av en flercellig organism i reproduktion. Utan döden skulle det inte bli något generationsskifte - en av de viktigaste förutsättningarna för den evolutionära processen.
Åldersrelaterade förändringar i åldringsprocessen består inte i alla fall i en minskning av organismens anpassningsförmåga. Hos människor och högre ryggradsdjur får man erfarenhet av livets process, förmågan att undvika potentiellt farliga situationer utvecklas. Immunförsvaret är också intressant i detta avseende. Även om dess effektivitet i allmänhet avtar efter att kroppen når mognad, på grund av det "immunologiska minnet" i förhållande till vissa infektioner, kan gamla individer vara mer skyddade än unga.
HYPOTES SOM FÖRKLARAR ÅLDRANDETS MEKANISMER
Gerontologi känner till minst 500 hypoteser som förklarar både grundorsaken och mekanismerna för åldrande av kroppen. De allra flesta av dem har inte klarat tidens tand och är av rent historiskt intresse. Dessa inkluderar i synnerhet hypoteser som kopplar samman åldrande med konsumtion av en speciell substans av cellkärnor, rädsla för döden, förlust av vissa icke-förnybara ämnen som kroppen tar emot vid tidpunkten för befruktning, självförgiftning med avfallsprodukter, och toxiciteten hos produkter som bildas under påverkan av tjocktarmens mikroflora. De hypoteser som idag är av vetenskapligt värde faller i en av två huvudriktningar.
Vissa författare betraktar åldrande som en stokastisk process av åldersrelaterad ackumulering av "misstag" som oundvikligen inträffar under normala livsprocesser, såväl som skador på biologiska mekanismer under påverkan av inre (spontana mutationer) eller externa (joniserande strålning) faktorer. Stokasticitet beror på den slumpmässiga karaktären hos förändringar i tid och lokalisering i kroppen. I olika versioner av hypoteserna i denna riktning är den primära rollen tilldelad olika intracellulära strukturer, på den primära skadan av vilka funktionella störningar på cell-, vävnads- och organnivåer beror. Först och främst är det den genetiska apparaten hos celler (hypotes om somatiska mutationer). Många forskare associerar de initiala förändringarna i en organisms åldrande med förändringar i strukturen och följaktligen i de fysikalisk-kemiska och biologiska egenskaperna hos makromolekyler: DNA, RNA, kromatinproteiner, cytoplasmatiska och nukleära proteiner och enzymer. Lipider urskiljs också cellmembran ofta måltavla av fria radikaler. Misslyckanden i receptorernas arbete, i synnerhet cellmembranen, stör effektiviteten av regleringsmekanismer, vilket leder till en obalans i vitala processer.
Den riktning som behandlas inkluderar också hypoteser som ser den grundläggande principen för åldrande i det ökande slitaget av strukturer med åldern i intervallet från makromolekyler till organismen som helhet, vilket i slutändan leder till ett tillstånd som är oförenligt med livet. Denna uppfattning är dock för enkel.
Kom ihåg att förekomsten och ackumuleringen av mutationsförändringar i DNA motarbetas av naturliga antimutationsmekanismer och de skadliga konsekvenserna av bildandet av fria radikaler
minska på grund av antioxidantmekanismernas funktion. Således, om "konceptet med slitage" av biologiska strukturer korrekt återspeglar essensen av åldrande, så är resultatet i form av en större eller mindre frekvens av senila förändringar i den ålder då dessa förändringar blir uppenbara hos olika människor en följd av överlagringen av destruktiva och skyddande processer. I detta fall inkluderar slitagehypotesen oundvikligen
faktorer som genetisk predisposition, tillstånd och till och med livsstil, som, som vi har sett, åldrandehastigheten beror på.
Den andra riktningen representeras av genetiska eller programhypoteser, enligt vilka åldrandeprocessen är under direkt genetisk kontroll. Denna kontroll, enligt vissa åsikter, utförs med hjälp av speciella gener. Enligt andra åsikter är det förknippat med närvaron av speciella genetiska program, vilket är fallet med andra stadier av ontogeni, såsom embryonala.
Till förmån för programmerat åldrande ges bevis, av vilka många redan har diskuterats i Sec. 8.6.1. Vanligtvis hänvisar de också till förekomsten i naturen av arter där förändringarna snabbt ökar efter reproduktion, vilket leder till att djuren dör. Typiskt exempel- Stillahavslax (sockeye lax, rosa lax) dör efter lek. Utlösningsmekanismen i detta fall är förknippad med en förändring i sättet för utsöndring av könshormoner, vilket bör betraktas som ett inslag i det genetiska programmet för den individuella utvecklingen av laxfiskar, vilket återspeglar deras ekologi och inte som universell mekanismåldrande.
Det är anmärkningsvärt att kastrerad rosa lax inte leker och lever 2-3 gånger längre. Det är under dessa extra år av livet som tecken på åldrande i celler och vävnader bör förväntas. Vissa programhypoteser bygger på antagandet att den biologiska klockan fungerar i kroppen, i enlighet med vilka åldersrelaterade förändringar inträffar. "Klockans" roll tillskrivs i synnerhet tymuskörteln, som slutar fungera när kroppen övergår till vuxen ålder. En annan kandidat är nervsystemet, särskilt några av dess avdelningar (hypothalamus, sympatiska nervsystemet), vars huvudsakliga funktionella element är i första hand åldrande nervceller. Låt oss anta att upphörandet av tymus fungerar vid en viss ålder, som utan tvekan är under genetisk kontroll, är en signal om början av åldrande av organismen. Detta innebär dock inte en genetisk kontroll av åldringsprocessen. I frånvaro av tymus försvagas immunologisk kontroll över autoimmuna processer. Men för att dessa processer ska starta behövs antingen mutanta lymfocyter (DNA-skada) eller proteiner med förändrad struktur och antigena egenskaper.
Gerontologi och geriatrik
Gerontologi (från grekiskan gerontos - gammal man) är en gren inom biologi och medicin som studerar mönstren för åldrande hos levande varelser, inklusive människor. Gerontologins huvudriktningar inkluderar studiet av de viktigaste orsakerna, mekanismerna och villkoren för åldrande, sökandet efter effektiva medel för att öka förväntad livslängd och förlänga perioden med aktiv arbetsförmåga.
Geriatrik (av grekiskan iatreia - behandling) är ett område inom klinisk medicin som studerar diagnos, behandling och förebyggande av sjukdomar hos äldre och senila.
- I kontakt med 0
- Google Plus 0
- OK 0
- Facebook 0






