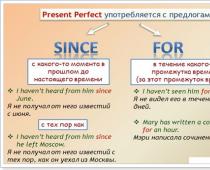
Allmänna egenskaper av typen Coelenterates.
Coelenterates är tvåskiktiga djur med radiell symmetri.
Symmetri. Kroppen av coelenterates har en huvudaxel, i ena änden av vilken det finns en munöppning. Flera symmetriaxlar passerar genom huvudaxeln, längs vilken djurets bihang och inre organ är belägna. Denna typ av symmetri kallas radiell .
Livsformer. De huvudsakliga livsformerna för coelenterates är polyper och maneter.
Kropp polyp allmänt cylindrisk, i ena änden finns en munöppning omgiven av ett varierande antal tentakler, och i den andra en sula. Polyper är vanligtvis stillasittande eller inaktiva. Polyper för det mesta bilda kolonier.
Kropp manet har utseendet av ett vanligt paraply eller klocka, på vars nedre, konkava sida det finns en munöppning. Det finns tentakler eller lober längs kanten av paraplyet och ibland runt munnen. Maneter leder som regel en aktiv livsstil och bildar inte kolonier.
Olika arter av coelenterates existerar antingen i form av endast en av dessa livsformer (maneter eller polyp), eller under hela deras liv livscykel gå igenom båda stadierna.
Taxonomi. I phylum Coelenterata finns det tre klasser:
Hydroid (hydra, obels, polypodium, sifonoforer);
Scyphoid (manetaurelia, cornerota, cyanea, havsgetingar);
Korall (svarta och röda koraller, acropora, svampar, havsanemoner, alcyonium).
Totalt finns det 9 000 moderna arter av coelenterater.
Kroppsmått coelenterates varierar kraftigt. Vissa typer av polyper i vuxen ålder överstiger inte några millimeter, medan vissa havsanemoner kan nå 1 meter i diameter. Hos maneter kan paraplyets diameter vara från 2 mm till 2 meter. Dessutom kan tentaklerna hos vissa maneter sträcka sig upp till 30 meter.
Rörelse. Polyper stillasittande. De kan böja sin kropp, dra ihop sig och röra sina tentakler. Hydras kan "gå" som mallarver (malarnas larver). Havsanemoner kan krypa långsamt på sulorna.
Manet aktivt röra sig genom att dra ihop paraplyet. Havsströmmar spelar också en viktig roll när de transporterar maneter över långa avstånd.
Kroppstruktur. Som redan nämnts är coelenterater tvåskiktsdjur. Deras kroppsvägg består av två celllager - ektoderm (extern) och endoderm (inre). Mellan dem är mesoglea - ett lager av strukturlös gelatinös substans. Den enda hålighet i kroppen av coelenterates - tarm, eller mag- .
Ectoderm presenteras som en enskiktsplatta, kubisk eller cylindrisk epitel . Utöver vanliga epitelceller inkluderar ektodermen epitelmuskulära celler vars bas är långsträckt längsgående riktning kontraktila (muskel)fiber. Hos vissa koraller separeras muskelfibrer från epitelet och ligger under det eller är nedsänkta i mesogleaskiktet och bildar ett oberoende muskelsystem. Mellan epitelceller finns interstitial celler som ger upphov till olika cellulära element i ektodermen. Ett karakteristiskt drag hos coelenterates är närvaron av så kallade tentakler i ektodermen stickande celler . Varje sådan cell innehåller en kapsel i vilken en spirallindad lång ihålig process sätts in - en stickande filament. Utanför cellen finns ett känsligt hår, när den är irriterad, visar sig den stickande tråden skarpt, rätar ut och genomborrar bytet eller fiendens kropp. Samtidigt hälls ett giftigt sekret ut ur kapseln, vilket orsakar förlamning hos små djur, såväl som en brännande känsla hos stora.
Endoderm. Epitelet som täcker maghålan består av flagellära celler. Några av dessa celler är epitelmuskulära Däremot är muskelprocesserna belägna i tvärriktningen och bildar tillsammans lager av ringformade fibrer. Ektodermala epitelceller kan bilda pseudopodier, med vars hjälp de fångar upp matpartiklar. Det finns även körtelceller.
Mesoglea. Hos polyper är mesogleaen dåligt utvecklad (med undantag för koraller), men hos maneter når detta lager en betydande tjocklek. Mesoglea innehåller ett antal ektodermala celler som deltar i bildningen av skelettet.
Skelettformationer. Endast polyper har ett skelett. I hydroidpolyper är kroppen täckt med en tunn kitinös theca - ett tätt membran som utför en skyddande funktion. De flesta typer av koraller har ett kalkhaltigt skelett, ibland ett kått. Skelettutvecklingen kan variera från individuella spikler utspridda i mesoglea till kraftfulla stenliknande strukturer av olika storlekar och former (i madrepore-koraller). Dessa skelett är ett derivat av ektoderm.
Bildandet av skelettet i koraller beror till stor del på närvaron av polyper i kroppen symbiotiska alger . Låt oss överväga kemiska reaktioner, som uppstår under bildningen av kalkskelettet. Utgångsämnena - kalciumjoner och koldioxid - finns i havsvatten i tillräcklig mängd.
Koldioxid, som löses i vatten, bildar mycket instabil kolsyra:
H 2 O + CO 2 ↔ H 2 CO 3, som omedelbart dissocierar till joner:
H2CO3 ↔ H+ + HCO3-.
När Ca-joner interagerar med HCO 3 bildas kalciumbikarbonat:
Ca ++ + 2 HCO 3 - ↔ Ca (HCO 3) 2. Detta ämne är lösligt i vatten, men det är inte starkt och förvandlas lätt till olösligt kalciumkarbonat:
Ca (HCO 3) 2 ↔ Ca CO 3 ↓ + H 2 O + CO 2.
Med ett överskott av CO 2 skiftar denna reaktion åt vänster och lösligt bikarbonat bildas. När CO 2 -koncentrationen minskar skiftar reaktionen åt höger och en kalkfällning faller ut.
Alger som lever i polyperkroppen avlägsnar ständigt koldioxid från vävnaderna i coelenterates för fotosyntesprocessen, vilket ständigt skapar en minskad koncentration av CO 2. Sådana förhållanden gynnar bildandet av olösligt kalciumkarbonat och konstruktionen av ett kraftfullt skelett av polyper.
Matsmältningssystemet och näring. Matsmältningssystemet representeras av maghålan. De flesta coelenterater är rovdjur. De för byten dödade eller bedövade av stickande celler med sina tentakler till munöppningen och sväljer det.
I hydroidpolyper ser maghålan ut som en enkel säck, som kommunicerar med miljö munöppning. Olika smådjur som kommer in i maghålan absorberas oftast av endodermceller ( intracellulär matsmältning). Större byten kan smältas av enzymer som utsöndras av körtelceller. Osmälta rester drivs ut genom munnen.
I korallpolyper är maghålan delad i längdriktningen av septa, vilket ökar arean av endodermen. Dessutom sticker det ektodermala svalget ut i matsmältningshålan hos koraller.
Som redan nämnts inleder revbyggande koraller ett symbiotiskt förhållande med en viss typ av encelliga alger som sätter sig i det endodermala lagret. Dessa växter, som tar emot koldioxid och metaboliska produkter från polypen, förser den med syre och ett antal organiska ämnen. Algerna själva smälts inte av polyperna. Under normala förhållanden tillåter en sådan symbios polyper att överleva under lång tid utan intag av organiska ämnen från miljön.
U manet Maghålan bildas vanligtvis av magsäcken, belägen i den centrala delen av paraplyet, radiella kanaler som sträcker sig från magen och en ringformad kanal som löper längs kanten av paraplyet. Det finns oftast 4 radiella kanaler i hydromedusae, och i scyphomedusae 16. Hela komplexet av kanaler bildar s.k. gastrovaskulära systemet .
Nervsystem. U polyper nervsystem diffus typ . Individuella nervceller som ligger vid basen av epitelet av ektodermen och endodermen är anslutna genom sina processer till ett nervnätverk. Munöppningen och basen av polyperna är omgivna av ett tätare nervnätverk.
U manet nervsystemet är mer koncentrerat än hos polyper, vilket är förknippat med en mobil livsstil.
U hydroid maneter kluster av nervceller finns längs kanten av paraplyet. Cellerna själva och deras processer bildar en dubbel nervring. Den yttre ringen utför sensoriska funktioner och den inre ringen utför motoriska funktioner.
U scyphoid maneter nervringen är mindre uttalad, men vid basen av rhopalia (marginala känselkroppar) finns kluster av nervceller som kan kallas ganglier.
Sinnesorgan. På grund av en stillasittande livsstil, polyper särskild inga sinnesorgan . Det finns bara enskilda känsliga (taktila) celler, som mestadels är belägna nära munöppningen.
U manet Det finns även sinnesceller, men det finns också speciella sinnesorgan – syn, balans och lukt.
Längs kanten av paraplyet finns synorgan - ögon , olika i struktur. Hos hydroidmaneter ligger ögonen ensamma, medan ögonen hos scyphoidmaneter ligger på rhopalia - känsliga marginalkroppar. Dessutom kan en ropalia bära flera ögon av varierande grad av komplexitet samtidigt.
På grund av sin aktiva livsstil har maneter utvecklats balansorgan - statocyster. De är en blåsa kantad från insidan med känsliga celler. Inuti bubblan finns en kalkhaltig kropp - en statolit. Beroende på manetens position i rymden irriterar statoliten ett visst område av vesikelväggen. Det finns andra typer av statocyststruktur. Dessutom kan statocyster upptäcka vibrationer i vatten, så de kan också kallas hörselorgan. Hos hydroidmaneter är balansorganen placerade längs kanten av paraplyet i mängden 4-80 olika typer.
Scyphoid maneter har också luktgropar är kemiska sinnesorgan.
Hos scyphoider är alla sinnesorgan belägna på 8 rhopalia - modifierade tentakler.
Andetag. Gasutbyte i coelenterater sker genom diffusion av syre och koldioxid. Hos stora arter (koraller) har svalget sifonoglyfer kantade med cilierade epitel. Celler utrustade med cilia levererar ständigt färskt vatten till djurets tarmhåla. Många polyper har, som redan nämnts, gått över till symbios med alger, som förser coelenteraten med syre och befriar dem från koldioxid.
Könsorgan. U polyper Det finns inga speciella könsorgan. Könsceller bildas antingen i ektodermen eller i endodermen. I det första fallet går könscellerna ut genom ett avbrott i ektodermen, i det andra går de först in i maghålan och sedan ut genom munnen. Bland polyperna finns hermafroditer (hydra) och tvåbo (koraller).
U manet , som nästan alltid är tvåbo, har könskörtlar.
U hydromedusa de bildas i ektodermen på undersidan av paraplyet under de radiella kanalerna, mindre ofta på den orala snabeln. Antalet könskörtlar motsvarar antalet radiella kanaler. Gameter frigörs genom bristningar i körtlarna.
U scyphoid maneter gonader av endodermalt ursprung. De bildas i fickorna i magen. Gameter kommer först in i maghålan och sedan in i miljön.
Fortplantning. Coelenterates reproducerar sig både asexuellt och sexuellt.
Asexuell fortplantning sker oftast genom gryende . Denna väg är typisk för polyper, men är sällsynt hos maneter. I enstaka polyper uppstår en knopp på kroppen, som gradvis bildar tentakler och en munöppning och sedan bryter av från moderns kropp. I koloniala hydroider och koraller separeras inte dotterindividen från modern, vilket leder till bildandet av kolonier.
Kolonial hydroidpolyper är inte kapabla till sexuell reproduktion, så de knoppar också av sexuella individer - maneter. Maneter bildas antingen på kolonins axel eller på speciella utväxter - blastostyles.
En annan metod för asexuell reproduktion är strobilation , när polypen i ett visst skede börjar snöra i tvärriktningen flera gånger och en liten manet bildas från varje del. Nästan hela kroppen av polypen spenderas på bildandet av maneter. Denna metod är typisk för scyphoid maneter.
Således finns det en förändring mellan polypoida asexuella och medusoida sexuella generationer. Samtidigt dominerar polypoidgenerationen i hydroider, medan medusoidgenerationen dominerar hos scyphoider. Koraller har inte en medusoid generation.
I ett antal hydroider separeras inte maneter från kolonin, och i vissa reduceras maneten till tillståndet av en "genitalsäck" - en sporosarca.
Mycket intressant sifonoforer , som representerar en enorm koloni bestående av organismer med olika strukturer. Varje koloni har en pneumatofor - en luftbubbla som stöder sifonoforen på vattenytan.
Sexuell fortplantning karakteristisk för alla maneter, alla koraller och vissa hydroidpolyper. Den sexuella processen involverar haploida celler - könsceller, som parar sig antingen i miljön eller i kroppen av coelenteratet. Ägget genomgår fullständig enhetlig krossning. Gastrulation av blastula sker oftast genom immigration, mer sällan genom intussusception. Därefter bildas en tvåskiktslarv - en planula, täckt med cilia och leder en aktiv livsstil. För stillasittande djur som koraller (som inte har en medusoid generation) är planula det enda spridningsstadiet. En polyp bildas alltid av planulan, som sedan knoppar av antingen bara polyper (koraller), eller polyper och maneter (hydroider), eller bara maneter (scyphoider). Sålunda sker utvecklingen av de allra flesta coelenterater med metamorfos. Ibland bildas en polyp omedelbart från ett ägg (till exempel i hydra).
Regeneration. Coelenterater har en hög förmåga att regenerera. Experiment för att studera detta fenomen i hydra utfördes redan 1740 av Tremblay. Det visade sig att djuret kan regenerera från 1/200 av delen.
Ursprung. Troligtvis var förfäderna till coelenterates frisimmande organismer som parenchymella , som beskrevs av I.I. Mechnikov. Dessa hypotetiska organismer saknade skelett och kunde därför inte ha bevarats som fossil.
De äldsta fynden av coelenterates - korallskelett - går tillbaka till den kambriska perioden (ca 600 miljoner år sedan). Samtidigt bevarades inte bara enskilda tryck utan även hela fossiliserade rev. Några få tryck av maneter och hydroider är också kända. Totalt är mer än 20 000 arter av fossila coelenterater kända.
Menande. I naturen deltar coelenterater, som är rovdjur och samtidigt mat för andra djur, i komplexa näringskedjor av marina biocenoser. Koraller är av stor geokemisk betydelse och bildar tjocka lager av kalkstenar. Under hela sin existens deltar koraller i bildandet av öar. Rev är unika biocenoser där ett stort antal djurarter lever.
Praktisk betydelse moderna coelenterates är liten.
Koraller (särskilt röda och svarta) används som smycken. De bryts i stora mängder främst med hantverksmetoder. På stora rev är det förbjudet att samla koraller.
Vissa maneter utgör en allvarlig fara för människor. I våra hav inkluderar dessa den lilla korsmaneten från Fjärran Östern, som lever i snår av marina växter, och den stora Svartahavskornetten, som ofta finns utanför kusten. Korsets gift är ibland dödligt. Den farligaste maneten, havsgetingen, lever utanför Australiens kust. Att röra vid detta djur orsakar svår smärta och chock. Många människor dog när de träffade henne.
I Kina och vissa andra länder äts speciellt beredda rhopile maneter. Det finns ett speciellt hantverk där.
Den mest typiska representanten för klassen är hydra(Fig. 7).
Den första personen som såg hydran var mikroskopets uppfinnare och den största naturforskaren på 1600- och 1700-talen. A. Levenguk (1632 – 1723).
När han tittade på vattenväxter såg han bland de små organismerna en märklig varelse med många "horn". Han observerade också tillväxten av knoppar på dess kropp, bildandet av tentakler i dem och separationen av det unga djuret från moderns kropp.
Hydra är en enkelpolyp i sötvatten med en avlång säckliknande kropp ca 1 cm lång Kroppen består av två lager av celler: yttre - ektoderm, och internt - endoderm, fodrar tarmhålan. De två lagren av celler är åtskilda av en tunn stödplatta - mesoglea. I den övre änden av hydras kropp finns en mun omgiven av en kronkrona med 6-12 tentakler. Med deras hjälp fångar hydra bytet och leder det in i munnen. I den nedre änden av kroppen finns en sula, med hjälp av vilken hydran är fäst vid undervattensobjekt.
Del ektoderm omfattar celler av olika typer: epitelmuskulära, stickande, mellanliggande, nervösa (fig. 8).
Epitelmuskelceller utgör grunden för ektodermen. Kontraktila fibrer i sina cellers processer ger rörelsen av tentaklerna och hela kroppen, som kan sträcka sig, dra ihop sig och gå som malfjärilars larver.
Ris. 7. Schematisk längsgående sektion av hydra: 1 - tentakel; 2 - mun; 3 - ektoderm; 4 - endoderm; 5 - mesoglea; 6 - tarmhålan; 7 - njure; 8 - manlig gonad; 9 – kvinnlig reproduktionskörtel.
Bland epitelmuskelcellerna är stickande celler lokaliserade ensamma eller i grupper. Det är särskilt många av dem på tentaklerna. Cellens ihåliga kapsel innehåller en spirallindad stickande filament. Ligger på cellens yttre yta känsligt hår, vars irritation (mekanisk eller kemisk) gör att sticktråden skjuter ut. De stickande cellerna används bara en gång och dör sedan.
För att ersätta de förbrukade stickande cellerna, såväl som andra typer av celler, utvecklas nya i ektodermen - från många små, snabbt förökande odifferentierade mellanceller. Tack vare deras närvaro har hydra en väl uttryckt förmåga att regenerera förlorade eller skadade celler och kroppsdelar.

Ris. 8. Hydrakroppsceller: A– epitelmuskulär ektodermcell; b– nervceller kopplade till varandra genom processer; V- två stickande celler (1 – i vila; 2 – urladdad).
Nervceller är belägna jämnt djupt i ektodermen; deras processer bildar ett nätverksliknande plexus - ett diffust nervsystem. Irritation från en cell överförs till andra nervceller och från dem till hud-muskelceller. Svaret på extern stimulering i hydran är en enkel obetingad reflex.
Således utför ektodermceller skyddande, motoriska och sensoriska funktioner.
Endodermen bildas av två typer av celler: körtelceller och matsmältningsceller. Körtelceller utsöndrar matsmältningsenzymer i tarmhålan. Matsmältningsceller liknar till strukturen de epitelmuskulära cellerna i ektodermen, men till skillnad från dem är de utrustade med en eller två flageller och kan bilda pseudopoder.
Följaktligen är endodermceller specialiserade på att utföra matsmältningsfunktioner.
Hydra – rovdjur djur. Med sina tentaklers stickande trådar slår den till små vattenlevande djur, förlamar och sväljer dem. I tarmhålan halvsmälts maten till ett mosigt tillstånd av enzymer som utsöndras av endodermens körtelceller. Små matpartiklar fångas sedan upp av rotationsrörelserna av flagellerna i matsmältningscellerna och fagocyteras av deras pseudopoder. Osmälta matrester tas bort genom munnen.
Således har hydra, som alla coelenterater, matsmältning blandad.
Fortplantning hydra förekommer under den varma årstiden asexuellt - genom knoppning. På hydrakroppen bildas först en liten tuberkel - en knopp, som är ett utsprång utåt från två lager av kroppen. Njuren ökar i storlek, tentakler och en munöppning bildas på den. Snart separeras den unga hydran från modern.
Med riklig näring reproducerar hydras genom att knoppas under den varma perioden på året. Med början av höstkylan börjar hydran att Sexuell fortplantning. Hydras av olika arter kan vara tvåbo och hermafrodit. Vissa mellanliggande ektodermceller differentierar till manliga och kvinnliga könsceller, som ackumuleras i den nedre eller mellersta delen av kroppen och kallas könskörtlar eller könskörtlar. I de utvecklande gonaderna ackumuleras ett stort antal mellanliggande, odifferentierade celler, från vilka både framtida könsceller och "näringsceller" bildas, på grund av vilka det framtida ägget ökar. I de första stadierna av äggutvecklingen förvandlas dessa celler till mobila amöboider. Snart börjar en av dem absorbera de andra och ökar avsevärt i storlek och når 1,5 mm i diameter. Denna stora amöboid, som plockar upp pseudopodia, blir rundad och blir ett ägg. Efter att den genomgått meios spricker gonadens vägg och ägget kommer ut, men förblir dock anslutet till hydrakroppen med en tunn plasmastjälk. Varje kvinnlig gonad producerar ett ägg.
Vid denna tidpunkt utvecklas spermier i testiklarna på andra hydras, som lämnar gonaden och simmar i vattnet. En av dem penetrerar ägget, varefter det omedelbart börjar separera. Det utvecklande embryot är täckt med två skal, vars yttre har täta kitinösa väggar och är ofta täckt med ryggar.
Skyddad av ett dubbelt skal - embryothecae– embryot övervintrar, medan vuxna hydror dör med början av kallt väder. På våren, inuti embryotheca, finns det redan en bildad liten hydra, som kommer ut genom en bristning i dess vägg.

Ris. 9. Schema för ett längdsnitt av en hydroid manet: Till vänster – en sektion i den radiella kanalens plan: 1 – munöppning; 2 - mage; 3 – orala tentakler; 4 - radiell kanal; 5 – segel; 6 – marginell tentakel; 7 - motorisk nervring; 8 - titthål; 9 - känslig nervring; 10 - gonad; till höger - en sektion mellan de radiella kanalerna: 11 - ektoderm, 12 - endoderm; 13 - mesoglea; 14 – ringkanal.
Mycket mer komplex hydroid maneter(Fig. 9). Externt ser hydromedusa ut som en genomskinlig skiva, paraply eller klocka. En oral snabel med en mun i änden hänger från paraplyets inre mitt. Kanterna på munnen kan vara släta eller utrustade med fyra mer eller mindre fransade munlober. Munnen leder till magen, som upptar hela håligheten i munsnabeln, fyra radiella kanaler sträcker sig från magen till paraplyets periferi. Vid kanten av paraplyet rinner de ut i en ringkanal. Kombinationen av magen och kanalerna kallas gastrovaskulära systemet. Längs kanten av hydromedusa-paraplyet finns tentakler och känselorgan. Tentaklerna används för att röra och fånga byten, de är tätt kantade med stickande celler.
Vissa hydromedusae har ljuskänsliga organ - ögon, som alltid är belägna vid basen av tentaklarna och är tydligt synliga på grund av sin mörka färg. Ögat består av två typer av celler - ljuskänslig och pigmenterad.Ögonen ser ut som fläckar eller gropar. I de mest komplexa ögonen är fossas hålighet fylld med ett genomskinligt ämne som fungerar som en lins.
Manetens rörelse utförs på grund av sammandragningen av muskelfibrer i kanten av paraplyet. Genom att trycka ut vatten ur paraplyhålan får maneten en jetpush och rör sig framåt med paraplyets ovansida. Att stärka den reaktiva förmågan uppnås på grund av närvaron på insidan av paraplyet av en ringformad utväxt, kallad ett segel, som smalnar av utgången från paraplyets hålighet.
Maneter är tvåbo, deras könskörtlar finns antingen i ektodermen av den orala snabeln eller i ektodermen av paraplyet under de radiella kanalerna. Här ligger de närmast de näringsämnen som behövs för utvecklingen av reproduktionsprodukter. Strukturen hos cellerna i ektodermen och endodermen hos maneter är densamma som hos polyper, men mesoglea är utan tvekan mer utvecklad. Den är rik på vatten och har en gelatinös natur, på grund av vilken hydromedusae är mycket genomskinliga; många, till och med ganska stora, maneter är svåra att se i vattnet. Mesoglea i paraplyet är särskilt starkt utvecklad.
Hydroider är en klass av djur som tillhör phylum Cnidarians (tillhör Coelenterates). Hydroider inkluderar mer än 2,5 tusen arter, inklusive sötvattenhydra. Men till skillnad från hydra, lever de flesta hydroider i haven och oceanerna och har två former i sin livscykel - en kolonial polyp och en manet. Alla hydroider har ett larvstadium från vilket en polyp utvecklas.
Både polypen och maneterna är rovdjur som fångar byten med sina tentakler.
Hydroidpolyper bildar kolonier som ett resultat av asexuell reproduktion genom knoppning. Till skillnad från ensamma hydras, i marina representanter, är dotterindivider inte separerade från sina mödrar. Deras tarmhåligheter kommunicerar med varandra, detta gör att mat som fångas av en polyp kan omfördelas i hela kolonin.
Hydroid maneter är små i storleken (inom 1 cm). De bildas på några polyper i kolonin, bryter sig loss från dem och börjar simma.
Maneten ser ut som en inverterad polyp. Dess kroppsform är dock inte cylindrisk, utan liknar ett paraply eller en klocka, det vill säga kroppsformen är mer tillplattad. En manet kan simma genom att dra ihop sitt paraply och släppa ut vatten från det, men oftare simmar hydroidmaneter passivt på grund av strömmen. Längst ner är munöppningen omgiven av tentakler. Till skillnad från polyper har maneter ett tjockare lager av mesoglea (det gelatinösa ämnet mellan ektodermen och endodermen).

Med hjälp av maneter upplever hydroider Sexuell fortplantning. Varje manet utvecklar könskörtlar underifrån. Hos vissa individer innehåller de ägg, hos andra innehåller de spermier; d.v.s. hydroidmaneter är tvåbodjur. När könscellerna mognar släpps de ut i vattnet där befruktning sker. Det händer att äggen inte lämnar maneten.
Den resulterande zygoten förvandlas till en larv täckt med flageller (mossmoppelarv). En sådan larv kan aktivt simma. Efter att ha sjunkit till botten ger den upphov till en hydroid polyp.
Hydras har inte ett manetstadium. Könscellerna mognar direkt på polypen. Det finns en åsikt att i evolutionsprocessen reducerades manetstadiet i dem.
Maneter tillåter hydroider att spridas över större områden.
Till skillnad från andra coelenterater har hydroider inga skiljeväggar i tarmhålan. Det vill säga de kan betraktas som mer primitiva coelenterater.
Underklass av hydroider (Hydroidea)
Kroppen av en hydroid polyp är cylindrisk eller äggformad, i sin ömma del är den vanligtvis utrustad med en stjälk, med vilken enkla polyper är fästa vid marken, vattenväxter, blötdjursskal, d.v.s. i allmänhet till undervattensobjekt.
I koloniala former förbinder en stjälk polypen med kolonin.
 I den övre änden av polypens kropp finns en munöppning omgiven av tentakler, som kan ordnas i vanliga kronkronor eller utan strikt ordning.
I den övre änden av polypens kropp finns en munöppning omgiven av tentakler, som kan ordnas i vanliga kronkronor eller utan strikt ordning.
Antalet tentakler kan variera mellan arter.
Till exempel har den gigantiska enkelhydroidpolypen Branchiocerianthus imperator, som når mer än 1 m i höjd, upp till 380 tentakler, medan den lilla monobrachium parasiticum bara har en. Oftare är varje polyp utrustad med 10-30 tentakler, som kan vara enkla eller utrustade med en stiftformad förtjockning i slutet.
Hos en art (Cladocoryne) grenar tentaklarna jämnt.
Det finns även polyper utan tentakler. Dessa inkluderar sötvattensmikrohydra (Microhydra). Polyper fångar mat med sina tentakler, och ensamma polyper, som sötvattenshydran, använder dem när de rör sig.
Den inre strukturen hos en hydroidpolyp är mycket enkel.
Det ser ut som en tvålagersväska. Utsidan av polypen är täckt med ett lager av ektodermceller, och dess tarmhåla (maghålan) är fodrad med endoderm.
Mellan lagren av ektoderm och endoderm i hydroidpolyper finns ett tunt lager av icke-cellulär substans - mesoglea. Ektodermen bildas av speciella epitel-muskelceller. En sådan cells kropp ser ut som ett mångfacetterat prisma; cellerna ligger nära, som en bikaka.
Den prismatiska delen utför den integumentära funktionen.
I sin inre ände har epitelcellerna en lång förlängning som sträcker sig upp och ner längs polyppens kropp. En tunn muskelfiber löper inuti denna process. Samma kontraktila processer sträcker sig från endodermceller, men de är belägna vinkelrätt mot ektodermala cellers processer.
Kombinationen av muskelfibrer från många ektodermala och endodermala celler gör att polypen och dess tentakler kan sträcka sig och dra ihop sig.
Stickande celler finns mellan epitel-muskelcellerna.
Ektodermen innehåller även speciella nervceller utrustade med långa tunna processer, som tillsammans bildar nätverksliknande nervplexus, något tätare i ändarna av tentaklarna och runt munnen.
I polyper som kan föröka sig sexuellt (till exempel hydra) utvecklas även könsceller i ektodermen, som ackumuleras i nedre eller mellersta delen av kroppen.
Dessa celler kallas gonader eller gonader. Slutligen finns här i epitelet mellan- eller reservceller från vilka epitelmuskulära, stickande, nerv- och könsceller av hydroidpolyper utvecklas.
 Ektodermen hos många typer av polyper utsöndrar ett tunt skal på utsidan - theca, bestående av kitin.
Ektodermen hos många typer av polyper utsöndrar ett tunt skal på utsidan - theca, bestående av kitin.
I hydroidpolyper fungerar detta membran som ett exoskelett, som fungerar som stöd och skydd.
Hos vissa arter bildar theca en blomkål runt polyperna - en hydrotheca, i vilken de dras in i händelse av fara.
Endodermen består av körtel- och epitelmuskulära celler. Körtelceller producerar matsmältningsjuicer (enzymer) som hjälper till att smälta maten.
Små djur som kommer in i polypens maghåla smälts och sönderdelas under påverkan av matsmältningsjuicer.
Hydroidpolyper. Strukturera. Metagenes
Endodermceller har 2-5 tunna flageller, som ständigt slingrar sig och flyttar innehållet i maghålan. Matpartiklar som befinner sig nära väggarna i maghålan fångas upp av pseudopodier som bildas på ytan av epitelmuskelceller. Ytterligare assimilering av mat sker inuti cellen, liknande vad som händer hos encelliga djur.
Matsmältningsprodukterna överförs till ektodermcellerna, separerade från endodermen med en tunn men tätt lager mesoglea, genom direkta kontakter av endoderm- och exodermceller, vars utväxter korsar mesogleaskiktet, tätt vidrör varandra.
I kolonier kommunicerar maghålorna hos individuella polyper med varandra och bildar en enda kolonihåla.
Föreläsning tillagd 2012-08-24 kl. 05:17:45
Klass Hydrozoa
Hydra. Obelia. Hydrans struktur. Hydroidpolyper
De lever i havet och sällan i sötvatten. Hydroider är de enklast organiserade coelenteraten: en maghåla utan septa, ett nervsystem utan ganglier, och könskörtlarna utvecklas i ektodermen. Bildar ofta kolonier. Många har ett generationsskifte i sin livscykel: sexuella (hydroida maneter) och asexuella (polyper) (se.
Coelenterater).
Hydra sp.(Fig. 1) - en enda sötvattenpolyp. Längden på hydrans kropp är cirka 1 cm, dess nedre del - sulan - tjänar till att fästa på substratet; på motsatt sida finns en munöppning, runt vilken 6-12 tentakler är placerade.
Liksom alla coelenterater är hydraceller arrangerade i två lager.
Det yttre skiktet kallas ektoderm, det inre skiktet kallas endoderm. Mellan dessa lager finns basalplattan. Följande typer av celler särskiljs i ektodermen: epitelmuskulär, stickande, nervös, mellanliggande (interstitiell). Alla andra ektodermceller kan bildas från små odifferentierade interstitiella celler, inklusive könsceller under reproduktionsperioden.
Vid basen av epitel-muskelcellerna finns muskelfibrer längs kroppens axel. När de drar ihop sig förkortas hydrans kropp. Nervcellerna är stjärnformade och placerade på basalmembranet. Förbundna med sina långa processer bildar de ett primitivt nervsystem av den diffusa typen. Svaret på irritation är reflexivt till sin natur.

ris. 1.
Hydra:
1 - mun, 2 - tunga, 3 - maghåla, 4 - ektoderm,
5 - endoderm, 6 - stickande celler, 7 - interstitiell
celler, 8 - epitelmuskulär ektodermcell,
9 - nervcell, 10 - epitelmuskulär
endodermcell, 11 - körtelcell.
Ektodermen innehåller tre typer av stickande celler: penetranter, volventer och glutinanter.
Den penetrerande cellen är päronformad, har ett känsligt hår - cnidocil, inuti cellen finns en stickande kapsel, som innehåller en spiralvriden stickande tråd.
Kapselhåligheten är fylld med giftig vätska. I änden av sticktråden finns tre taggar. Att röra vid cnidocil orsakar att en stickande tråd frigörs. I det här fallet genomborras ryggarna först i offrets kropp, sedan injiceras giftet från den stickande kapseln genom trådkanalen.
Giftet har en smärtsam och förlamande effekt.
De andra två typerna av stickande celler utför den ytterligare funktionen att hålla kvar byte. Volventer skjuter fångstrådar som trasslar in offrets kropp. Glutinanter släpper ut klibbiga trådar. Efter att trådarna skjutit ut dör de stickande cellerna. Nya celler bildas från mellanliggande celler.
Hydra livnär sig på små djur: kräftdjur, insektslarver, fiskyngel, etc.
Bytet, förlamat och immobiliserat av stickande celler, leds in i maghålan. Matsmältning av mat är hålrum och intracellulära, osmälta rester utsöndras genom munnen.
Maghålan är kantad med endodermceller: epitelmuskulära och glandulära. Vid basen av de epitel-muskulära cellerna i endodermen finns muskelfibrer belägna i tvärriktningen i förhållande till kroppens axel; när de drar ihop sig, smalnar kroppen av hydran.
Området av epitelmuskelcellen som vetter mot maghålan bär från 1 till 3 flageller och kan bilda pseudopoder för att fånga matpartiklar. Förutom epitel-muskulära celler finns körtelceller som utsöndrar matsmältningsenzymer i tarmhålan.

ris. 2.
Klass Hydrozoa
Hydra spirande:
1 - moderns individ,
2 - dotterindivid (knopp).
Hydra reproducerar sig asexuellt (knoppande) och sexuellt.
Asexuell reproduktion sker under vår-sommarsäsongen. Knopparna bildas vanligtvis i kroppens mittområden (fig. 2). Efter en tid separeras unga hydras från moderns kropp och börjar leva ett självständigt liv.
Sexuell reproduktion sker på hösten. Under sexuell reproduktion utvecklas könsceller i ektodermen.
Spermier bildas i områden av kroppen nära munnen, ägg - närmare sulan. Hydras kan vara antingen tvåbo eller hermafroditiska.
Efter befruktning täcks zygoten med täta hinnor och ett ägg bildas.
Hydran dör, och en ny hydra utvecklas från ägget följande vår. Direkt utveckling utan larver.
Hydra har en hög förmåga att regenerera. Detta djur kan återhämta sig även från en liten avskuren del av kroppen.
Interstitialceller är ansvariga för regenereringsprocesser. Hydras vitala aktivitet och regenerering studerades först av R. Tremblay.
Obelia sp.- en koloni av marina hydroidpolyper (fig. 3). Kolonin ser ut som en buske och består av individer av två typer: hydranthus och blastostyles.
Ektodermen hos medlemmarna i kolonin utsöndrar ett skelett organiskt skal - peridermen, som utför funktionerna stöd och skydd.
De flesta av kolonins individer är brandposter. Strukturen hos en brandpost liknar den hos en hydra.
Till skillnad från hydra: 1) munnen är belägen på munstjälken, 2) munstjälken är omgiven av många tentakler, 3) maghålan fortsätter i den gemensamma "stammen" i kolonin.
Mat som fångas av en polyp distribueras bland medlemmar av en koloni genom de grenade kanalerna i den gemensamma matsmältningshålan.

ris. 3. Obelias livscykel:
1 - koloni av polyper, 2 - hydroid maneter,
3 - ägg, 4 - planula,
5 - ung polyp med en njure.
Blastostilen har formen av en stjälk och har ingen mun eller tentakler.
Manetknopp från blastostyle. Maneter bryter sig loss från blastostilen, flyter i vattenpelaren och växer. Formen på hydroidmaneten kan jämföras med formen på ett paraply. Mellan ektodermen och endodermen finns ett gelatinöst lager - mesoglea. På den konkava sidan av kroppen, i mitten, på munstjälken finns en mun.
Många tentakler hänger längs kanten av paraplyet och tjänar till att fånga byten (små kräftdjur, larver av ryggradslösa djur och fiskar). Antalet tentakler är en multipel av fyra. Mat från munnen kommer in i magen, fyra raka radiella kanaler sträcker sig från magen och omger kanten på manetens paraply.
Metoden för maneternas rörelse är "reaktiv"; detta underlättas av vecket av ektoderm längs kanten av paraplyet, kallat "seglet". Nervsystemet är av diffus typ, men det finns kluster av nervceller längs kanten av paraplyet.
Fyra gonader bildas i ektodermen på den konkava ytan av kroppen under de radiella kanalerna.
Könsceller bildas i gonaderna.
Från det befruktade ägget utvecklas en parenkymlarv, motsvarande en liknande svamplarv. Parenkymulen förvandlas sedan till en planulalarv i två lager.
Planulan, efter att ha simmat med hjälp av cilia, lägger sig till botten och förvandlas till en ny polyp. Denna polyp bildar en ny koloni genom knoppning.
Obelias livscykel kännetecknas av växling mellan asexuella och sexuella generationer. Den asexuella generationen representeras av polyper, den sexuella generationen av maneter.
Beskrivning av andra klasser av typen Coelenterates.
Marina, mer sällan sötvattensdjur som leder en fast livsstil eller simmar i vatten. Bifogade blanketter kallas polyper, flytande - manet.
Dubbelt lager djur, deras kropp består av två cellulära lager: yttre - ektoderm och internt - endoderm. Endoderm former tarm, eller maghålan. Maghålan kommunicerar med omgivningen genom en öppning som fungerar som oral Och anal. Mellan ektoderm och endoderm är mesoglea. Hos polyper bildar mesoglea en stödjande platta, medan den hos maneter bildar ett tjockt gelatinartat lager.
Ektodermceller utför skyddande och motoriska funktioner. Ektodermen innehåller speciella stickande celler som tjänar till försvar och attack. Endodermceller kantar maghålan och utför huvudsakligen en matsmältningsfunktion. Matsmältning intracellulärt Och hålighet.
Andning sker genom hela kroppens yta.
Nervsystem tankspridd, eller diffus, typ. Tillgängliga taktil känslighet, och hos maneter är de, på grund av sin simlivsstil, ljusuppfattande "ögon" Och balansorgan.
Coelenterates har radiell, eller radiell, symmetri.
Asexuell fortplantning gryende. Könsorgan presenteras könskörtlar. Befruktningen är extern. Vissa representanter kännetecknas av alternerande asexuella (polyp) och sexuella (maneter) generationer i livscykeln.
Typen av coelenterater inkluderar följande klasser: Hydrozoaner, Scyphoid maneter, korallpolyper.
Klass Hydrozoa
Sötvattenhydra
EN KORT BESKRIVNING AV
|
Livsmiljö |
Tvåskiktsdjur i sötvatten. Led en kopplad livsstil |
|
Utseende |
Säckform upp till 1,5 cm Radiell symmetri. Munnen i den främre änden av kroppen är omgiven av tentakler, sulan är den bakre änden av kroppen, för fastsättning |
|
Kroppsskydd |
Ektoderm - yttre lager, endoderm - inre lager, mesoglea - mellanlager |
|
Kroppshålighet |
Det finns ingen kroppshåla. Det finns bara en tarmhåla |
|
Matsmältningssystemet |
Blint stängd tarmhåla. Munöppning för matintag och för utdrivning av osmält matrester. Matsmältning intrakavitär och intracellulär |
|
utsöndringsystemet |
Ektodermceller |
|
Nervsystem |
Nervceller av stjärntyp. Diffusa nervsystemet |
|
Sinnesorgan |
Ej utvecklad |
|
Andningssystem |
Ingen. Andas genom hela kroppens yta |
|
Fortplantning |
Asexuell - genom knoppning. Hermafroditer. Korsbefruktning. |
GENERELLA EGENSKAPER
Denna klass inkluderar små former av coelenterater. Polyper Och manet som tillhör denna klass kallas hydroid.
Strukturera . Hydrans kropp är avlång dubbellagerspåse, fäst vid basen, eller enda till substratet (fig. 1). Yttre lager - ektoderm, inre lager - endoderm. Det finns utrymme mellan lagren - mesoglea.
I den fria änden av kroppen finns oral kon, omgiven av en kant av 6-12 tentakler. Ligger på munkäglan mun, anställd och anus. Hela kroppsytan är täckt ektoderm, huvudsakligen bestående av cylindrisk eller kubiska epitelceller. Deras bas förlängs uppåt och nedåt, längs kroppens längsgående axel, till en lång process. Processens cytoplasma skiljer sig som kontraktila fibrer, i samband med detta spelar processen muskulös roll. De cylindriska delarna av cellerna bildas enkelskikts epitel. Således utför cellerna en dubbel funktion - omslag Och motor och kallas epitelmuskulära. Med samtidig sammandragning av alla muskelprocesser förkortas hydrakroppen. Mellan epitel-muskelcellerna finns små mellanliggande celler som deltar i bildandet stickande Och könsceller, och även i processen regeneration- återställande av förlorade kroppsdelar eller organ. Direkt under epitelet finns stjärnformade nervceller. Förbundna av sina processer bildar nervceller nervsystemet tankspridd, eller diffus, typ. Av särskild betydelse i ektodermen är stickande celler, eller kapslar, tjänar för attack och försvar.
Endoderm kantar helheten mag-, eller matsmältningshålan. Grunden för endodermceller är epitelmuskulära matsmältningsceller. Dessa cellers muskelprocesser, till skillnad från ektodermala, är belägna på tvären med avseende på kroppens längdaxel. När de drar ihop sig smalnar hydrans kropp ihop och blir tunnare. Endodermala celler inkluderar körtelceller, utsöndrar matsmältningsenzymer i maghålan, och celler med fagocytisk aktivitet. De senare är kapabla att fånga matpartiklar med hjälp av rörelsen av 1-3 flageller och bildandet av pseudopodier. Således kombinerar hydra två typer av matsmältning: intracellulärt Och kavitär.
Ris. 1.Strukturen av sötvattenhydra: a - längsgående sektion; b - tvärsnitt; c - tvåskiktskropp; d - epitelmuskelcell; d - tentakel med kasserade stickande trådar; f, g - stickande celler; 1 - tentakler; 2 - testis; 3 - spermier; 4 - maghålan; 5 - spirande ung hydra; 6 - stödplatta; 7 - endoderm; 8 - ektoderm; 9 - ägg i olika utvecklingsstadier; 10 - stickande celler; 11 - munöppning; 12 - sula
Mesoglea presenteras i form av en tunn strukturlös platta - basalmembran.
Asexuell fortplantning. Ungefär i nivå med mitten av hydras kropp finns en sk spirande bälte, där det bildas då och då knopp, av vilken en ny individ sedan bildas. Efter bildandet av munnen och tentaklerna är knoppen vid basen lös, faller till botten och börjar existera självständigt. Denna metod för asexuell reproduktion kallas gryende.
Sexuell fortplantning . När kallt väder närmar sig börjar hydrorna att reproducera sig sexuellt. Mellanceller i ektodermen kan omvandlas direkt till ägg eller genom multipel division - in spermier. Mellanceller som bildar ägg ligger närmare basen av hydran och de som bildar spermier - till munöppningen. Äggen befruktas i moderns kropp på hösten och omges av ett tätt skal, då dör modern, och äggen förblir vilande till våren. På våren utvecklas en ny individ från dem. Hydras tvåbo, men de träffas och hermafroditiska slag.
Marina hydroidpolyper
De flesta marina hydroidpolyper bildar kolonier. Kolonier har oftast formen av ett träd eller en buske. Stamgrenarna, grenarna bildar separata kolonier - brandposter. Maghålorna i alla brandposter kommunicerar med varandra, så mat som fångas upp av en brandpost distribueras över hela kolonin. I marina hydroidpolyper bildar det ektodermala epitelet ett speciellt membran - strömmande, vilket ger hela kolonin större stabilitet.
Marina hydroidpolyper reproducerar sig bara asexuellt- gryende. Sexuell fortplantning fullgöra sexuella individer- manet, som bildas på en polyp genom knoppning och övergång till en frisimande livsstil. Maneter har dock samma struktur som polyper
det finns också skillnader (fig. 2, 3). Maneternas kropp karakteriseras stark utveckling av mesoglea som innehåller en stor mängd vatten. Nervsystemet är också mycket mer komplext. Hos maneter, längs kanten av paraplyet, en solid nervring. Det finns sinnesorgan: ögon Och statocyster (jämviktsorgan). Manet tvåbo. Sexkörtlar ligger på undersidan av paraplyet mellan ektodermen och mesoglea. Befruktning och utveckling av ägg sker i den yttre miljön. Ägg utvecklas till larver parenkymula, sedan den andra larven - planula, som flyter fritt en tid, sedan sjunker till botten och ger upphov till en polyp. En ny koloni bildas därefter från polypen, och cykeln upprepas. Således består livet av hydroidpolyper av två generationer. En generation- polyper, leda en stillasittande livsstil och fortplanta sig asexuellt. Andra generationen - maneter, leda en frisimmande livsstil och fortplanta sig sexuellt. Det vill säga i hydroidpolyper förekommer det generationsväxling.

Ris. 2.Strukturen av en hydroid polyp (A) och en hydroid manet (B), inverterad med munnen öppnande uppåt: 1 - mun; 2 - tentakler; 3 - maghålan; 4 - mesoglea; 5 - radiell kanal; 6 - segla

Ris. 3Schema för strukturen hos en hydroid manet: 1 - mun; 2 - oral stjälk med gonad (3); 4 - radiella kanaler; 5 - ringkanal; 6 - tentakler; 7 - ögon; 8 - segla
Klass Scyphoid maneter
Denna klass inkluderar manet, lever bara i haven. De är större än hydroidmaneter och deras struktur är mer komplex (Fig. 4). Munnen slutar i ett svalg, och maghålan är uppdelad i kammare. Den ringformiga kanalen, som löper längs kroppens kant, förenar kanalerna som sträcker sig från magen och bildar gastrovaskulära systemet. Kluster av nervceller visas i formen ganglier. Könsceller bildas i könskörtlar- könskörtlar lokaliserade i endodermen. Utvecklingen fortskrider med generationsväxling (fig. 5).

Ris. 4.Schema för strukturen hos en scyphoid manet: 1 - munlober; 2 - munöppning; 3 - tentakler; 4 - ringkanal; 5 - radiell kanal; 6 - gonad; 7 - magtrådar; 8 - mage; 9 - ektoderm; 10 - mesoglea; 11 - endoderm

Ris. 5.Utveckling av scyphoid maneter: 1 - ägg; 2 - planula; 3 - scyhistoma; 4 - spirande scyhistoma; 5 - strobilation; 6 - eter; 7 - vuxna maneter
Klass korallpolyper
Korallpolyper har bara en livsform - polyp. De har inte generationsväxling. Marina, ensamma, mestadels koloniala djur. Korallpolyper skiljer sig från andra klasser genom närvaron av ett hårt kalkhaltigt skelett, såväl som muskelfibrer i ektodermen och endodermen, vilket gör att de kan ändra kroppens form.
När vi vandrar längs havsstranden ser vi ofta åsar av grönaktiga, bruna eller bruna trassliga klumpar av hårda trådar som kastas ut av vågorna. Väldigt få människor vet att en betydande del av detta "havsgräs" inte är av växt, utan av animaliskt ursprung. Alla som varit på havet har förstås sett att alla stenar, högar och andra undervattensföremål är övervuxna med någon sorts ömtåliga buskar som vrider sig i vågorna. Om du samlar sådana buskar och tittar på dem i mikroskop, kan du tillsammans med riktiga alger se något väldigt speciellt. Här framför oss finns en brun segmenterad gren med rosa klumpar i ändarna. Till en början är de rosa klumparna orörliga, men så fort de står tysta i några minuter börjar de röra sig, sträcka ut sig i längd, tar formen av en liten kanna med en krona av tentakler i den övre delen av kroppen . Dessa är hydroidpolyper eudendrium(Eudendrium), lever i våra norra hav, i Svarta havet och i haven vidare Långt österut. I närheten finns en annan, också segmenterad, men lättare gren. Polyperna på den är också rosa, men formade som en spindel. Tentaklarna sitter på polypkroppen utan någon ordning, och var och en är utrustad i slutet med ett litet huvud - ett kluster av stickande celler. Polypernas rörelser är långsamma, de böjer ibland sin kropp, ibland svajar långsamt från sida till sida, men oftare sitter de orörliga, med sina tentakler brett isär - de ligger och väntar på byten. På vissa polyper kan man se knoppar eller unga maneter under utveckling. Vuxna maneter klämmer kraftigt och löser upp sitt paraply, den tunna tråden som förbinder maneten med polypen går av och maneten simmar iväg med ryck. Dessa är polyper Corine(Cogune) och deras maneter. De lever också i både arktiska och tempererade hav.
Och här är en annan buske, polyperna på den sitter inuti genomskinliga klockor. Utåt är de väldigt lika Eudendrium-polyper, men beter sig helt annorlunda. Så fort du lätt rör vid polypen med änden av nålen dras den snabbt tillbaka till djupet av sitt skyddande skal - en klocka. På samma buske kan du också hitta maneter: de, som polyper, är gömda inuti ett genomskinligt skyddande skal. Maneter sitter tätt på en tunn tentakellös polyp. Detta är en hydroidkoloni obelia(Obelia).
Nu när vi kan skilja hydroider från alger bör vi vara uppmärksamma på den fjäderliknande kolonin aglaofeni(Aglaofeni). Hos denna art, som är mycket vanlig i vår Svartahavsregion, sitter matpolyperna på en gren i en rad. Var och en är innesluten i en blomkål, hydrotheca, och omgiven av tre skyddande polyper.
Aglaophenia producerar inte frisimmande maneter, och underutvecklade individer av medusoidgenerationen är gömda inuti en mycket komplex formation - en korg (en modifierad gren av kolonin).
Kolonier av hydroider bosätter sig oftast på grunt djup - från kustzonen till 200-250 m och föredrar stenig jord eller fäster vid olika trä- och metallföremål. De växer ofta mycket tätt på undervattensdelarna av fartyg och täcker dem med en lurvig "pälsrock". I dessa fall orsakar hydroider betydande skada på sjöfarten, eftersom en sådan "päls" kraftigt minskar fartygets hastighet. Det finns många fall där hydroider, som sätter sig inuti rören i ett marint vattenförsörjningssystem, nästan helt stängde deras lumen och förhindrade tillförsel av vatten. Det är ganska svårt att bekämpa hydroider, eftersom dessa djur är opretentiösa och utvecklas ganska bra, verkar det i ogynnsamma förhållanden. Dessutom kännetecknas de av snabb tillväxt - buskar 5-7 cm höga växer på en månad. För att rensa dem från botten av skeppet måste du lägga det i torrdocka. Här rensas fartyget från övervuxna hydroider, polychaetes, mossor, sjöekollon och andra beväxta djur.
I Nyligen Särskilda giftiga färger började användas, de undervattensdelarna av fartyget belagda med dem är utsatta för nedsmutsning i mycket mindre utsträckning.
Hydroider som sätter sig i kustzonen är inte alls rädda för bränningen. I många av dem skyddas polyperna från slag av en skelettkopp - theca; på kolonier som växer i själva surfzonen är thecae alltid mycket tjockare än de av samma art som lever djupare, där de brytande vågorna inte känns (fig. 159).

I andra hydroider från surfzonen har kolonier långa, mycket flexibla stammar och grenar, eller så är de uppdelade i segment. Sådana kolonier slingrar sig tillsammans med vågorna och går därför inte sönder eller går sönder.
På stora djup lever speciella hydroider som inte liknar kustarter. Kolonier i form av fiskben eller fjäder dominerar här, många ser ut som träd, och det finns arter som liknar en borste. De når en höjd av 15-20 cm och täcker havsbotten med tät skog. Maskar, blötdjur, kräftdjur och tagghudingar lever i snåren av hydroider. Många av dem, till exempel havsgeters kräftdjur, finner en tillflykt bland hydroider, andra, som havsspindlar (flerledda), gömmer sig inte bara i sina snår utan livnär sig också på hydropolyper.
Om du flyttar ett finmaskigt nät runt hydroidboplatser eller, ännu hellre, använder ett speciellt, så kallat planktonnät, kommer du bland massan av små kräftdjur och larver av olika andra ryggradslösa djur att stöta på hydroidmaneter. De flesta arter av hydromedusa är inte särskilt stora djur, de når sällan mer än 10 cm i paraplydiameter, vanligtvis är storleken på en hydromedusa 2-3 cm, och ofta bara 1 - 2 mm. Hydroid maneter är mycket genomskinliga. Du kommer inte ens att märka maneter som fångas och placeras i glasfat direkt: bara de vitaktiga trådarna i kanalerna och den orala snabeln är synliga. Bara genom att titta noga kan du lägga märke till paraplyets konturer.
Tittar på en hydroidkoloni Korine(Sogupe), vi har redan sett nykläckta små maneter av denna art. En fullformad manet har ett klockformat paraply 1-8 cm högt, fyra tentakler och en lång, maskliknande munsnabel. Med skarpa sammandragningar av paraplyet rör sig maneten snabbt i ett horisontellt plan eller stiger uppåt. Den sjunker sakta ner under påverkan av gravitationen, frusen i vattnet med lösa tentakler. Marina planktoniska kräftdjur, som utgör maneternas huvudföda, gör ständigt vertikala rörelser: under dagen störtar de ner i djupet och på natten stiger de till ytan. De sjunker ner i djupare, lugna vattenlager även under vågor. Maneter rör sig ständigt efter dem; två sinnen hjälper dem att förfölja sitt byte - känsel och syn. I lugnt vatten drar manetens paraply ihop sig rytmiskt hela tiden och lyfter upp djuret till ytan. Så fort maneten börjar känna vattenrörelsen som orsakas av vågorna, slutar paraplyet att dra ihop sig och det sjunker sakta ner i djupet. Den upptäcker ljus med hjälp av ögonen som finns vid basen av tentaklerna. För starkt ljus verkar på det som spänning - paraplyet slutar dra ihop sig och djuret störtar ner i mörkare djup. Dessa enkla reflexer hjälper maneterna att förfölja byten och fly från katastrofal spänning.
Som nämnts ovan livnär sig Corine-maneten på planktoniska organismer, främst copepoder. Ögonen på en manet är inte så perfekta att den kan se sitt byte, den fångar det blint. Dess tentakler kan sträcka sig mycket och överstiga paraplyets höjd tiotals gånger. Hela ytan av tentakeln är prickad med många stickande celler. Så fort ett kräftdjur eller något annat litet planktondjur vidrör tentakeln påverkas det omedelbart av stickande celler.
Samtidigt drar tentakeln snabbt ihop sig och drar bytet till munnen. Den långa snabeln sträcker sig i riktning mot bytet. Om ett större kräftdjur fångas, flätar maneten ihop det inte med en, utan med två, tre eller alla fyra tentakler.
Maneter med platt paraply och många tentakler fångar sitt byte på ett helt annat sätt, t.ex. tiaropsis(Tiaropsis) är en hydromedusa storleken på ett tvåkopekmynt, mycket vanlig i våra norra hav. Längs kanterna på paraplyet finns det upp till 300 tunna tentakler. En vilande manet har tentakler vida åtskilda och täcker ett betydande område. När paraplyet drar ihop sig verkar maneterna sopa bort kräftdjuren med dem och trycka dem mot mitten av undersidan av paraplyet (se bild 160). Thiaropsis mynning är bred, utrustad med fyra stora fransade blad, med vilka maneten fångar de anpassade kräftdjuren.

Trots sin lilla storlek är hydroidmaneter väldigt glupska. De äter mycket kräftdjur och anses därför vara skadliga djur - konkurrenter till plankätande fiskar. Maneter behöver riklig mat för att utveckla reproduktiva produkter. Medan de simmar sprider de ett stort antal ägg i havet, vilket sedan ger upphov till polypoida generationen av hydroider.
Ovan kallade vi coelenterates typiska invånare i havet. Detta gäller för 9 000 arter som tillhör denna typ, men ungefär ett och ett halvt till två dussin arter av coelenterater lever i sötvatten och finns inte längre i haven. Tydligen flyttade deras förfäder till sötvatten för länge sedan.
Det är mycket karakteristiskt att alla dessa former av både sötvattens- och bräckvattenbassänger endast avser hydroid klass och till och med bara till en av honom underklass - hydroidea(Hydroidea).
Bland alla andra coelenterater observeras ingen förkärlek för vatten med låg salthalt.
Till de mest typiska invånarna i sötvatten överlag klot, som ofta bildar mycket täta populationer, omfattar flera arter hydr, komponenter hydra truppen(Hydrida).
FÄRSKVATTENHYDRA
I varje grupp av djurriket finns representanter älskade av zoologer, som de använder som huvudobjekt när de beskriver djurens utveckling och struktur och på vilka de utför många experiment inom fysiologi. I filumen Coelenterates är ett sådant klassiskt föremål hydra. Detta är förståeligt. Hydra är lätt att hitta i naturen och relativt lätt att hålla i laboratoriet. De förökar sig snabbt, och därför kan massamaterial erhållas på kort tid. Hydra är en typisk representant för coelenterates, som står vid basen av det evolutionära trädet av flercelliga organismer. Därför används den för att klargöra alla frågor som rör studiet av anatomi, reflexer och beteende hos lägre flercelliga organismer. Detta i sin tur hjälper till att förstå ursprunget för djur av högre ordning och utvecklingen av deras fysiologiska processer. Dessutom fungerar hydra som ett utmärkt objekt för utveckling av sådana allmänna biologiska problem som regenerering, asexuell reproduktion, matsmältning, axiell fysiologisk gradient och mycket mer. Allt detta gör det till ett oumbärligt djur för båda utbildningsprocess- från gymnasium upp till senior år på universitetet och i vetenskapligt laboratorium, där problemen med modern biologi och medicin i deras olika grenar löses.
Den första personen som såg hydran var mikroskopets uppfinnare och den största naturforskaren på 1600- och 1700-talen. Anton Levenguk.

När man tittade på vattenväxter såg Leeuwenhoek bland andra små organismer ett konstigt djur med många "horn". Han observerade också tillväxten av knoppar på dess kropp, bildandet av tentakler i dem och separationen av det unga djuret från moderns kropp. Leeuwenhoek avbildade en hydra med två njurar och ritade också spetsen av dess tentakel med stickande kapslar, som han såg den under sitt mikroskop.
Leeuwenhoeks upptäckt väckte dock nästan ingen uppmärksamhet från hans samtida. Bara 40 år senare blev de intresserade av hydra i samband med den extraordinära upptäckten av den unge läraren Trambley. Medan han studerade föga kända vattenlevande djur på sin fritid upptäckte Tremblay en varelse som liknade både ett djur och en växt. För att bestämma dess natur skar Tremblay varelsen på mitten. Den regenerativa förmågan hos lägre djur var fortfarande nästan okända vid den tiden, och man trodde att endast växter kunde återställa förlorade delar. Till Tremblays förvåning växte en hel hydra från varje halva, båda rörde sig, grep byte, vilket betyder att det inte var en växt. Möjligheten att förvandla en bit av en hydras kropp till ett helt djur hyllades som en betydande upptäckt inom livsvetenskapen, och Tremblay påbörjade en djup och seriös studie av hydran. 1744 publicerade han boken "Memoirs on the History of a Kind of Freshwater Polyps with Arms in the Form of Horns." Boken beskrev i detalj hydrans struktur, dess beteende (rörelser, fånga bytesdjur), reproduktion genom knoppning och vissa aspekter av fysiologi. För att testa sina antaganden utförde Tremblay en serie experiment med hydra, vilket lade grunden för en ny vetenskap om experimentell zoologi.
Trots den tidens ofullkomlighet i optiken och zoologins svaga utveckling skrevs Tremblays bok på en så hög nivå vetenskaplig nivå, som inte har förlorat sin betydelse till denna dag, och teckningar från denna bok finns i många läroböcker om zoologi.
Numera uppgår den vetenskapliga litteraturen om hydra till många hundra artiklar och böcker, men ändå upptar hydra forskarnas sinnen än i dag. Ett litet primitivt djur fungerar som en prövsten för dem, där många frågor löses modern vetenskap om livet.
Om du samlar vattenväxter från kustdelen av en sjö eller flod och placerar dem i ett akvarium med rent vatten, kommer du snart att se hydror på dem. Till en början är de nästan osynliga. Störda djur drar ihop sig kraftigt, deras tentakler drar ihop sig. Men efter en tid börjar hydrans kropp sträcka sig, dess tentakler förlängs. Nu syns hydran tydligt. Formen på dess kropp är rörformad, i den främre änden finns en munöppning omgiven av en kronkrona på 5-12 tentakler. Omedelbart under tentaklarna har de flesta arter av hydra en liten avsmalning, en hals, som skiljer "huvudet" från kroppen. Den bakre änden av hydran är avsmalnad till en mer eller mindre lång stjälk, eller stjälk, med en sula i änden (hos vissa arter uttrycks inte stjälken). I mitten av sulan finns ett hål, den så kallade aborala poren. Hydras maghåla är solid, det finns inga skiljeväggar i den, tentaklarna är ihåliga, liknar handskarnas fingrar.
Hydras kroppsvägg, liksom alla coelenterates, består av två lager av celler, deras fina struktur har redan beskrivits ovan, och därför kommer vi här att uppehålla oss vid endast en egenskap hos cellerna i hydrans kropp, som har hittills studerats fullständigt endast i detta objekt och har inte hittats i andra coelenterater.
Strukturen av ektodermen (och endodermen) i olika delar av hydras kropp är ojämlik. I huvudänden är alltså ektodermcellerna mindre än på kroppen; det finns färre stickande och mellanliggande celler, men en skarp gräns mellan integumentet av "huvudet" och kroppen kan inte dras, eftersom förändringen i ektoderm från kroppen till "huvudet" sker mycket gradvis. Hydrasulans ektoderm består av stora körtelceller, vid korsningen mellan sulan och stjälken förloras integumentära cellers körtelkaraktär gradvis. Detsamma kan sägas om endodermceller Matsmältningsprocesser sker i den mellersta delen av hydras kropp, här har dess endoderm ett stort antal matsmältningskörtelceller, och de epitel-muskulära cellerna i endodermen i den mellersta delen av kroppen bildas många pseudopodier. I huvuddelen av maghålan, i stjälken och i tentaklerna sker ingen matsmältning. I dessa delar av kroppen ser ektodermen ut som foderepitel, nästan utan matsmältningskörtelceller. Återigen kan en skarp gräns mellan cellerna i matsmältningsdelen av maghålan å ena sidan och sådana celler i "huvudet", stjälken och tentaklarna å andra sidan inte dras.
Trots skillnaden i strukturen hos cellskikten i olika delar av hydras kropp, är alla dess celler inte på strikt definierade permanenta platser, utan rör sig ständigt och deras rörelse är strikt regelbunden.
Genom att använda hydras höga förmåga att läka sår kan du göra detta intressant upplevelse. De tar två hydras av samma storlek och en av dem är målad med någon form av intravital färg, det vill säga ett färgämne som penetrerar hydrans vävnader utan att döda den. Vanligtvis används en svag vattenlösning av noll blau sulfat för detta, som färgar hydravävnaden i Blå färg. Efter detta genomgår hydrorna en operation: var och en av dem skärs i tre delar i tvärriktningen. Sedan fästs huvudet och nedre ändarna av det omålade provet på den mellersta delen av den "blåa" hydran. Skivorna växer snabbt ihop, och vi får en experimentell hydra med blått bälte mitt på kroppen. Strax efter operationen kan man observera hur det blå bandet sprider sig i två riktningar - mot huvudändan och stjälken. I det här fallet är det inte färgen som rör sig över hydrans kropp, utan själva cellerna. Skikten av ektoderm och endoderm tycks "flyta" från mitten av kroppen till dess ändar, medan naturen hos deras ingående celler gradvis förändras (se fig. 162).

I den mellersta delen av hydras kropp förökar sig cellerna mest intensivt, och härifrån rör de sig i två motsatta riktningar. Således förnyas cellsammansättningen ständigt, även om djuret utåt förblir nästan oförändrat. Denna egenskap hos hydran är mycket viktig när man löser frågor om dess regenerativa förmåga och för att bedöma data om förväntad livslängd.
Hydra är ett typiskt sötvattensdjur, endast i mycket sällsynta fall har hydrar hittats i svagt salthaltiga vattendrag, till exempel i Finska viken Östersjön, och i vissa bräckvattensjöar, om salthalten i dem inte översteg 0,5 %. Hydras lever i sjöar, floder, bäckar, dammar och till och med diken om vattnet är tillräckligt rent och innehåller en stor mängd löst syre. Hydras stannar vanligtvis nära kusten, på grunda platser, eftersom de älskar ljus. När du håller hydror i ett akvarium flyttar de sig alltid till den upplysta sidan.
Hydras är stillasittande djur, för det mesta sitter de på ett ställe, med sulorna fästa på en gren av en vattenväxt, en sten etc. Hydrans favoritställning i lugnt tillstånd är att hänga upp och ner, med lätt åtskilda tentakler hänger ner.
Hydra fäster på substratet tack vare det klibbiga sekretet från körtelcellerna i sulans ektoderm och använder även sulan som sugkopp. Hydran håller mycket stadigt och är ofta lättare att riva än att separera från underlaget. Om du tittar på en sittande hydra länge kan du se att dess kropp sakta svajar hela tiden, vilket beskriver en cirkel med dess främre ände. Hydra kan godtyckligt mycket snabbt lämna platsen där den sitter. Samtidigt öppnar den tydligen den aborala poren som ligger i mitten av sulan, och sugverkan stannar. Ibland kan du se hydran "gå". Först böjer den kroppen mot underlaget och stärker sig på den med hjälp av tentakler, sedan drar den upp den bakre änden och stärker den på en ny plats. Efter det första "steget" tar han det andra, och så vidare, tills han stannar på en ny plats.

Således rör sig hydran relativt snabbt, men det finns en annan, mycket långsammare, rörelsemetod - glidning på sulan. Med kraften från sulans muskler rör sig hydran knappt märkbart från plats till plats. Det tar väldigt lång tid att märka ett djurs rörelse. Hydras kan simma i vattenpelaren under en tid. Efter att ha lossnat från underlaget och spridit sina tentakler brett, faller hydran mycket långsamt till botten; den kan bilda en liten gasbubbla på sulan, som bär djuret uppåt. Hydras tar dock sällan till dessa rörelsemetoder.
Hydra är ett glupskt rovdjur, det livnär sig på ciliater, planktoniska kräftdjur, oligochaete maskar och attackerar också fiskyngel. Hydras ligger och väntar på sitt byte, hänger på någon kvist eller stjälk av en vattenväxt, och sprider sina tentakler brett och gör ständigt cirkulära letande rörelser. Så fort en av hydrans tentakler vidrör offret, rusar de återstående tentaklarna mot den och förlamar djuret med stickande celler. Nu finns det inga spår av hydrans långsamhet, den agerar snabbt och "bestämt". Bytet dras till munnen av tentakler och sväljs snabbt. Hydra sväljer små djur hela. Om bytet är något större än själva hydran kan det också svälja det. Samtidigt öppnas rovdjurets mun vida, och kroppens väggar sträcks kraftigt. Om bytet inte passar helt in i maghålan, sväljer hydran bara ena änden av det, och trycker offret djupare och djupare när det smälts. En välmatad hydra krymper något och dess tentakler drar ihop sig.
I maghålan, där matsmältningsprocesser precis har börjat, är omgivningens reaktion något alkalisk, och i matsmältningsvakuolerna i endodermen, där matsmältningen slutar, är den något sur. Hydra kan metabolisera fetter, proteiner och animaliska kolhydrater (glykogen). Stärkelse och cellulosa, som är av vegetabiliskt ursprung, absorberas inte av hydra. Osmälta matrester drivs ut genom munnen.
Hydras reproducerar sig på två sätt: vegetativt och sexuellt. Vegetativ reproduktion i hydras är av spirande natur. Knopparna visas i den nedre delen av hydrans kropp ovanför stjälken, efterföljande knoppar är något högre än de tidigare, ibland sitter de på motsatta sidor av hydrans kropp, ibland är de ordnade i en spiral (uppträdesordningen och knopparnas placering beror på typen av hydra). Samtidigt utvecklas 1-3, sällan fler, knoppar på hydrans kropp, men hydras med 8 eller fler knoppar har observerats.

I de första stadierna framträder njuren som en knappt märkbar konisk tuberkel, sedan sträcker den ut sig och antar en mer eller mindre cylindrisk form. I den yttre änden av knoppen uppträder tentaklernas rudiment, till en början ser de ut som korta trubbiga utväxter, men gradvis sträcker de ut sig och stickande celler utvecklas på dem. Slutligen tunnar den nedre delen av njurkroppen ut till en stjälk, och en munöppning bryter ut mellan tentaklarna. Den unga hydran förblir fortfarande ansluten till moderns kropp under en tid, ibland lägger den till och med knopparna för nästa generation. Separationen av knoppande hydras sker i samma sekvens som knopparna visas. Den unga hydran är något mindre i storlek än modern och har ett ofullständigt antal tentakler. De saknade tentaklarna dyker upp senare.
Efter riklig knoppning är moderhydran utmattad och inga knoppar visas på den på en tid.
Vissa forskare har också observerat uppdelningen av hydras, men denna metod för reproduktion bör tydligen klassificeras som onormala (patologiska) processer. Uppdelning i hydra uppstår efter skada på dess kropp och kan förklaras av den höga regenerativa förmågan hos detta djur.
Med riklig näring under hela den varma perioden på året, reproducerar hydras genom knoppning, de börjar sexuell reproduktion med början av hösten. De flesta arter av hydra är tvåbo, men det finns också hermafroditer, det vill säga de där både manliga och kvinnliga könsceller utvecklas på en individ.

Gonader bildas i ektodermen och ser ut som små tuberkler, kottar eller runda kroppar. Ordningen av utseende och arten av placeringen av gonaderna är samma som njurarna. Varje kvinnlig gonad producerar ett ägg.
I de utvecklande gonaderna ackumuleras ett stort antal mellanliggande, odifferentierade celler, från vilka både framtida könsceller och "näringsceller" bildas, på grund av vilka det framtida ägget ökar. I de första stadierna av äggutvecklingen får de mellanliggande cellerna karaktären av mobila amöboider. Snart börjar en av dem absorbera de andra och ökar avsevärt i storlek och når 1,5 mm i diameter. Efter detta tar den stora amöboiden upp sin pseudopodia och dess konturer blir rundade. Efter detta sker två mognadsdelningar, under vilka cellen delas i två ojämna delar, och på utsidan av ägget återstår två små så kallade reduktionskroppar - celler separerade från ägget till följd av delning. Under den första mognadsdelningen halveras antalet äggkromosomer. Det mogna ägget kommer ut från gonaden genom en lucka i dess vägg, men förblir ansluten till hydrakroppen med hjälp av en tunn protoplasmatisk stjälk.
Vid denna tidpunkt utvecklas spermier i testiklarna hos andra hydras, som lämnar gonaden och flyter i vattnet, en av dem penetrerar ägget, varefter krossningen omedelbart börjar.
Medan cellerna i det utvecklande embryot delar sig är utsidan täckt av två membran, vars yttre har ganska tjocka kitinoida väggar och ofta täckt med ryggar. I detta tillstånd övervintrar embryot under skydd av det dubbla skalet, embryotheca. (Vuxna hydras dör med början av kallt väder.) På våren, inuti embryotheca, finns det redan en nästan bildad liten hydra, som lämnar sitt vinterskal genom en bristning i dess vägg.
För närvarande är ett dussin arter av hydras kända som lever i sötvatten på kontinenter och många öar. De olika typerna av hydror skiljer sig väldigt lite från varandra. En av arterna kännetecknas av en ljusgrön färg, vilket beror på närvaron av symbiotiska alger i kroppen av dessa djur - zoochlorella. Bland våra hydras den mest kända stjälkad eller brun hydra(Hydra oligactis) och stamlös, eller - vanlig, hydra(Hydra vulgaris).
Hur beter sig hydran i sin omgivning, hur uppfattar den irritationer och reagerar på dem?
Liksom de flesta andra coelenterater, reagerar hydra på alla ogynnsamma irritationer genom att dra ihop kroppen. Om kärlet som hydrorna sitter i skakas lätt, då kommer några av djuren att dra ihop sig omedelbart, på andra kommer en sådan chock inte att ha någon effekt alls, några av hydrorna kommer bara att dra åt tentaklerna något. Detta gör att graden av reaktion på irritation i hydras är mycket individuell. Hydran saknar helt förmågan att "komma ihåg": du kan sticka den med en tunn stift i timmar, men efter varje sammandragning sträcker den sig igen i samma riktning. Om injektionerna är mycket frekventa, slutar hydran att svara på dem.
Även om hydror inte har speciella organ för att känna av ljus, reagerar de definitivt på ljus. Hydrans främre ände är mest känslig för ljusstrålar, medan dess stjälk nästan inte uppfattar ljusstrålar. Om du skuggar hela den gröna hydran kommer den att krympa på 15-30 sekunder, men om du skuggar en huvudlös hydra eller skuggar bara stammen på en hel hydra, så kommer den att krympa först efter 6-12 minuter. Hydras kan urskilja riktningen för ljusflödet och röra sig mot dess källa. Hastigheten för rörelsen av hydras mot ljuskällan är mycket låg. I ett av experimenten placerades 50 gröna och lika många bruna hydras i ett kärl på ett avstånd av 20 cm från glasväggen genom vilken ljuset föll. De gröna hydrorna var de första som rörde sig mot ljuset; efter 4 timmar nådde 8 av dem akvariets lätta vägg, efter 5 timmar fanns det redan 21 av dem och efter 6 timmar - 44. Vid det här laget kom de första 7 bruna hydrorna dit. I allmänhet visade det sig att bruna hydror var sämre i ljuset, först efter 10 timmar samlades 39 bruna hydror nära ljusväggen. De återstående försöksdjuren var fortfarande på väg vid det här laget.
Hydras förmåga att röra sig mot en ljuskälla eller helt enkelt flytta till lättare områden i poolen är mycket viktig för dessa djur. Hydras livnär sig huvudsakligen på planktoniska kräftdjur - cyklop och daphnia, och dessa kräftdjur vistas alltid på ljusa och väluppvärmda platser vid solen. Således närmar sig hydrar sitt byte när de går mot ljuset.
För en forskare som studerar lägre organismers reaktioner på ljus öppnar hydror upp det bredaste aktivitetsfältet. Experiment kan göras för att fastställa hur känsliga djur är för svaga eller omvänt mycket starka ljuskällor. Det visade sig att hydras inte reagerar alls på för svagt ljus. Mycket starkt ljus gör att hydran flyttar in i skuggade områden och kan till och med döda djuret. Experiment genomfördes för att fastställa hur känslig hydran är för förändringar i ljusintensitet, hur den beter sig mellan två ljuskällor och om den särskiljer enskilda delar av spektrumet. I ett av experimenten målades akvariets vägg i alla färger i spektrat, med gröna hydror som samlades i den blåvioletta regionen och bruna i den blågröna regionen. Detta innebär att hydras skiljer färg, och deras olika typer har olika "smak" för det.
Hydras (förutom grönt) behöver inte ljus för normal funktion. Om du matar dem bra, lever de bra i mörker. Den gröna hydran, i vars kropp de symbiotiska algen zoochlorella lever, mår dåligt även med överflöd av mat i mörkret och drar ihop sig kraftigt.
På hydras är det möjligt att utföra experiment på effekterna av olika typer av skadlig strålning på kroppen. Således visade det sig att bruna hydror dör efter bara en minuts belysning med ultravioletta strålar. Den gröna hydran visade sig vara mer resistent mot dessa strålar - den dör först vid den 5-6:e minuten av bestrålning.
Experiment på effekten av röntgenstrålar på hydra är mycket intressanta. Små doser av röntgenstrålar orsakar ökad knoppning i hydras. Bestrålade hydras, jämfört med icke-bestrålade, producerar ungefär 2,5 gånger fler avkommor under samma period. Ökning av stråldosen orsakar undertryckande av reproduktion; om hydran får för stor dos röntgenstrålar dör de strax efter. Det är viktigt att notera att låga doser av strålning ökar hydras regenerativa förmåga.
När hydra utsattes för radioaktiv strålning erhölls ett helt ovanligt resultat. Det är välkänt att djur inte känner radioaktiva strålar på något sätt och därför, om de kommer in i sin zon, kan de få en dödlig dos och dö. Den gröna hydran, som reagerar på radiumstrålning, försöker flytta sig bort från sin källa.
Från exemplen ovan är det tydligt att sådana experiment med hydrer, som att studera inverkan av olika miljöfaktorer på dem, inte är tomt kul, inte vetenskap för vetenskapens skull, utan en allvarlig och mycket viktig fråga, vars resultat kan ge mycket betydelsefulla praktiska slutsatser.
Naturligtvis genomfördes en studie av påverkan av temperatur, koncentration på hydra koldioxid, syre, samt ett antal gifter, mediciner, etc.
Hydra visade sig vara ett mycket bekvämt föremål för att utföra ett antal experimentell forskning om att studera fenomenet regenerering hos djur.
Som har nämnts många gånger, återställer hydra lätt förlorade kroppsdelar. Ett djur som skärs på mitten ersätter snart de saknade delarna. Men det blir oklart: varför växer alltid ett "huvud" med tentakler längst fram i segmentet och en stjälk baktill? Vilka lagar reglerar återvinningsprocesser? Det är ganska troligt att några av dessa lagar kan vara gemensamma för både hydra och mer välorganiserade djur. När du väl känner till dem kan du göra det viktiga slutsatser, tillämplig även på medicin.
Det är mycket enkelt att utföra operationer på hydras, du behöver inga bedövningsmedel eller komplicerade kirurgiska instrument. All utrustning i ”operationsrummet” består av en nål inbäddad med ett öga i ett trähandtag, en vass ögonskalpell, en liten sax och tunna glasrör. De första experimenten för att fastställa hydras regenerativa förmågor utfördes för mer än 200 år sedan av Tremblay. Denna noggranna forskare observerade hur hela djur dök upp från de längsgående och tvärgående halvorna av hydras. Sedan började han göra längsgående snitt och såg att stjälkar bildades från flikarna i polypens nedre del och "huvuden" bildades av flikarna i dess övre del. Genom att upprepade gånger operera på en av experimentpolyperna fick Tremblay en sjuhövdad polyp. Efter att ha skurit av alla sju "huvuden" för honom började Tremblay vänta på resultaten och såg snart att i stället för varje avskuret "huvud" hade ett nytt dykt upp. Den sjuhövdade polypen, i vilken de avskurna "huvudena" växer ut igen, var som två ärtor i en balja som den mytiska varelsen - den lerniska hydran, dödad av den store hjälten antikens Grekland Herkules. Sedan dess har sötvattenpolypen behållit namnet hydra.
Längs vägen konstaterade Tremblay att hydran återställs inte bara från halvor utan också från mycket små delar av kroppen. Det har nu konstaterats att även från 1/200 av en hydras kropp kan en hel polyp utvecklas. Men det visade sig senare att den regenererande förmågan hos sådana små bitar från olika delar av hydrans kropp inte är densamma. Området på sulan eller stjälken återställs till en hel hydra mycket långsammare än området från den mellersta delen av kroppen. Detta faktum förblev dock oförklarat under lång tid.
De inre krafterna som reglerar och styr processerna för normal regenerering avslöjades mycket senare av den berömda amerikanske fysiologen Child. Child konstaterade att ett antal lägre djur har en uttalad fysiologisk polaritet i sina kroppar. Således, under påverkan av giftiga ämnen, dör cellerna på djurets kropp och förstörs i en mycket specifik sekvens, nämligen från framsidan till baksidan (i Hydra, från "huvudet" till "sulan"). Därför är celler som finns i olika delar av kroppen fysiologiskt ojämlika. Skillnaden mellan dem ligger i många andra manifestationer av deras fysiologi, inklusive effekten på att utveckla unga celler på platsen för skadan.
Den gradvisa förändringen av cellers fysiologiska aktivitet från en pol till den andra (längs kroppsaxeln) kallas den axiella fysiologiska gradienten.
Nu blir det klart varför delarna som skärs från hydrasulan mycket långsamt återställer hypostomen och tentaklarna - cellerna som bildar dem är fysiologiskt mycket långt från cellerna som bildar "huvudet". Den axiella gradienten spelar en mycket viktig roll vid regenerering, men även andra faktorer har en märkbar inverkan på denna process. Under regenerering är närvaron på den regenererande delen av en utvecklande njure eller en artificiellt planterad bit vävnad från en annan del av djurets kropp, särskilt från dess främre del, mycket viktig. Med hög fysiologisk aktivitet påverkar de utvecklande njurarna eller "huvudcellerna" på ett visst sätt tillväxten av regenererande celler och underordnar deras utveckling deras inflytande. Sådana grupper av celler eller organ som gör sina egna justeringar av verkan av den axiella gradienten kallas organisatörer. Förtydligande av dessa egenskaper hos regenerering hjälpte till att förstå många oklara frågor i utvecklingen av djurorganismen.
I största centrum fysiologi - i institutet skapat av akademiker Pavlov i Koltushi finns ett monument över en hund. De flesta av de lagar som anges i Pavlovs läror upptäcktes under experiment på hundar. Kanske förtjänar den lilla sötvattenpolypen samma monument.
FÄRSKVATTENMANET
År 1880 dök plötsligt maneter upp i en pool av tropiska växter på London Botanical Society. Två zoologer Lankester och en stor expert på coelenterates Olmen (A1man) rapporterade denna upptäckt på sidorna i tidskriften Nechur (Nature). Maneterna var mycket små, den största av dem nådde knappt 2 cm i paraplydiameter, men deras utseende upphetsade dåtidens zoologer: innan dess hade de inte ens föreställt sig att sötvattensmaneter kunde existera. Maneter ansågs vara typiska invånare i havet. Inte långt innan detta hade den magnifika sydamerikanska vattenväxten Victoria Regia planterats i poolen, så det föreslogs att maneterna fördes till London tillsammans med plantmaterial från Amazonas. Efter en tid försvann maneterna från poolen lika mystiskt som de hade dykt upp. De upptäcktes igen bara fem år senare, också i London, men i en annan pool med samma tropiska växt. 1901 dök dessa maneter upp i Lyon (Frankrike), också i en växthuspool med Victoria Regia. Sedan började de hittas i München, Washington, St. Petersburg och Moskva. Maneter hittades antingen i bassängerna i botaniska trädgårdar eller i akvarier med tropiska fiskar. Till akvarieälskares förvåning fick de plötsligt nya husdjur. Små maneter (ofta bara 1 - 2 mm i paraplydiameter) dök plötsligt upp i stort antal i ett akvarium där det inte hade funnits några dagen innan. Under flera dagar kunde man observera hur maneter ryckigt rörde sig i vattnet och ivrigt åt små kräftdjur. Men en vacker dag när han tittade in i sitt akvarium hittade ägaren bara fisk i det, det fanns inga maneter där.
Vid denna tidpunkt beskrevs sötvattensmaneten i detalj i speciell zoologisk litteratur. Det visade sig att hon tillhör hydroid klass. De ringde henne kraspedakustoy(Craspedacusta). De minsta maneterna har ett halvklotformat paraply, 4 radiella kanaler och 8 tentakler. När maneten växer blir formen på dess paraply plattare och antalet tentakler ökar.

Mogna maneter når 2 cm i diameter och bär ett brett segel längs kanten av paraplyet och cirka 400 tunna tentakler fodrade med stickande celler. Den orala snabeln är tetraedrisk, med en korsformad munöppning, kanterna på munnen är något vikta. Vid den punkt där de radiella kanalerna avgår från den orala snabeln utvecklas 4 gonader. Maneter är mycket genomskinliga, deras mesoglea är färglös och deras tentakler, radiella kanaler, orala snabel och könskörtlar är vitaktiga eller krämfärgade.
Denna manet önskade zoologer svår gåta. Om vi håller med om att den hamnar i växthus tillsammans med växter från tropikerna, hur överlever den då transporter? Victoria regia transporterades från Amazonas stränder i form av frön eller rhizomer. Delika maneter, som av misstag fångas tillsammans med rhizomer, måste utan tvekan dö under den långa resan över havet. Men även om vi antar att maneten trots uttorkning kan överleva, hur kommer den då in i exotiska fiskälskares små akvarier?
Snart började maneter hittas i naturliga vattendrag. Första gången fångades hon i Yangtzefloden i Kina, sedan i Tyskland och sedan i USA. Men både i naturliga och konstgjorda reservoarer var upptäckter mycket sällsynta och alltid oväntade: till exempel upptäcktes maneter en gång i lagringsanläggningarna i Washingtons vattenförsörjningssystem.

Observationer av maneten har fastställt att den knoppar från små tentakellösa polyper som kallas mikrohydras(Mikrohydra). Dessa polyper hittades redan 1884 i samma pooler i London där maneter fångades, men då föreställde sig ingen ett samband mellan dessa två så olika varelser. Microhydra-polyper är synliga för blotta ögat som vita prickar mot bakgrund av gröna blad från vattenväxter som de brukar bosätta sig på. Deras höjd överstiger vanligtvis inte 0,5-1 mm, kroppsformen liknar en kägla: kroppen är i form av en flaska, och på en kort hals sitter ett sfäriskt "huvud" med en mun i mitten. Huvudet är tätt packat med stickande celler, det finns inga tentakler. Polyper bildar ibland primitiva kolonier på 2-7 individer. Microhydra reproducerar genom knoppning och bildar liknande tentakellösa polyper. Då och då separeras en grupp celler formad som en liten mask från ena sidan av polypens kropp. Sådana grupper av celler kallas frustler. Frustula kan slingra sig, krypa längs botten och klättra upp på vattenväxter, här förvandlas den till en ung mikrohydra.
En gång kunde jag observera hur en manet började utvecklas från en knopp på kroppen av en mikrohydra; när hon separerade från polypen och började simma var det lätt att känna igen henne som en ung craspedakusta. Det var också möjligt att följa utvecklingen av Kraspedakusta-ägg. Till en början bildas en maskliknande larv från ägget, utan flimmerhår och mycket lik microhydra frustula. Efter en period av krypning längs substratet fäster larven på det och förvandlas till en tentakellös polyp. Sålunda konstaterades att maneten craspedacusta och microhydrapolypen tillhör samma art av coelenterates, men till dess olika generationer.
Experimenten visade att under generationsväxlingen i denna art av hydroider är det extremt stort inflytande påverkas av miljöförhållanden. Knoppning av maneter på polyper sker endast vid en vattentemperatur på minst 26-33°C, och knoppning av polyper och separation av frustula - vid en temperatur av 12-20°C. Efter detta blev det klart att artens existens kunde upprätthållas under lång tid på grund av reproduktionen av polyper. Varken akvarister eller botaniker i växthus uppmärksammar små, orörliga mikrohydror, eftersom de är nästan osynliga för blotta ögat, och det är mycket svårt att hitta dem i naturen. Polyper kan leva länge i ett akvarium och när temperaturen stiger uppstår medusformade knoppar i alla polyper och de skiljer maneterna åt. Craspedacust-maneter är rörliga och kan ses i vattnet med blotta ögat. Nu blir det tydligt varför de nästan alltid hittades i pooler med tropiska växter och fiskar: dessa pooler värmdes upp på konstgjord väg. Bara en sak är oklart: levde maneter alltid i Europa eller fördes de dit? (Polyper kan kanske tåla viss uttorkning och en lång resa under ogynnsamma förhållanden.) Och var är hemlandet för microhydra craspedacusta?
Det är ganska svårt att svara på denna fråga. Sedan den första upptäckten av maneter i London har över 100 fall av deras närvaro i olika delar av världen beskrivits. Här kort beskrivning artens utbredning. I Sovjetunionen är deras livsmiljö Lyubov-reservoaren nära Tula, Donfloden, Lake Karayazi nära Tbilisi (på en höjd av nästan 2000 m över havet), Kurafloden och konstgjorda reservoarer i Gamla Bukhara. Dessutom har maneter och polyper upprepade gånger dykt upp i akvarier av amatörfiskodlare och vid universitet i Moskva och Leningrad. Utanför vårt land hittades denna art i nästan alla europeiska länder, i Indien, Kina och Japan, i Australien, norra och Sydamerika. Det är nu omöjligt att ange var dess hemland är och vart det fördes.
På senare tid fick denna art av coelenterater igen zoologer att tänka. Nu, när fördelningen, livsstilen, strukturen hos polyper och maneter verkade vara väl studerade, upptäcktes det plötsligt att polyper av två släkten kan utvecklas från Craspedakus-ägg - de tentakellösa som beskrivits ovan och de med tentakler. Båda typerna av polyper bildar frustula. Tentakelpolyper bildar genom knoppning liknande och icke-tentakelpolyper, de kan inte knoppa från maneter. Tentakellösa polyper bildar liknande polyper och maneter, men kan inte knoppa polyper utrustade med tentakler. Båda formerna av polyper bildas från frustula. Tentakler polyper har hittills bara upptäckts två gånger: 1960 i Ungern och 1964 i ett akvarium Leningrad universitet. Tillstånden som orsakade deras utseende är fortfarande oklara. Indiens floder och de stora sjöarna i Afrika är hem för ytterligare två arter av sötvattensmaneter, nära släktingar till Craspedakusta. En välkänd manet från den afrikanska Tanganyikasjön, kallad limnocnida(Limnocnida tanganjice).
URSPRUNG TILL FÄRSKVATTENKOELENTARITETER
Bland sådana hydroider är det först och främst nödvändigt att säga om Cordylophora.

Cordylophora bildar små ömtåliga kolonier i form av buskar upp till 10 cm höga Polyper sitter i ändarna av grenarna och har en spindelform. Varje polyp har 12-15 tentakler, som sitter i ingen strikt ordning i mitten av kroppen. Cordylophora har inte frisimmande maneter, individer av medusoidgenerationen är knutna till kolonin.
Denna art upptäcktes först av akademiker vid Ryska akademin P. S. Pallas 1771 i den norra delen av Kaspiska havet, varför cordylophora och kallas Caspian (Cordylophora caspia). Dess utbredning är dock inte alls begränsad till denna bassäng, den lever i Östersjön, Svarta och Azovska havet och finns också längs hela Europas Atlantkust och vid mynningen av alla större floder i Asien, Amerika och Australien. Denna art bosätter sig endast i mycket avsaltade områden i havet och lever på grunda djup, vanligtvis inte djupare än 20 m.
Namnet som Pallas gav Cordylophora - Kaspiska havet - har också sin egen betydelse. Faktum är att Cordylophoras hemland är Kaspiska havet. Först i mitten av förra seklet trängde cordylophora genom Volga- och Mariinsky-systemen in i Östersjön, där den på grund av sin låga salthalt (0,8%) fann sitt andra hem. Cordylophora är en tillväxtorganism; den lägger sig på alla fasta undervattensobjekt, både stationära och rörliga. Ytterligare hjälp vid vidarebosättning gavs av otaliga fartyg som flockades från alla sidor till Östersjön. När de återvände hem tog de från Östersjön på sin botten en objuden gäst, en "gränsintränger".
Men hur kom frilevande coelenterater in i sötvattenförekomster? Kunde de inte använda mynningen av floder som rinner ut i havet för detta? Naturligtvis kan de, men de kommer att behöva övervinna två hinder. En av dem är en minskning av salthalten. Endast arter som tål mycket betydande avsaltning kan komma in i floder.
Bland typiska marina invånare finns de för vilka även den minsta minskningen av saltprocenten i havsvatten har en skadlig effekt. Dessa inkluderar nästan alla korallpolyper, scyphoid maneter och de flesta hydroider. Men vissa hydroider kan fortfarande existera även med viss avsaltning. Av de coelenterates som nämns i denna bok är Corine en euryhalin. Denna art kan leva både i vatten med normal oceanisk salthalt och i avsaltade hav, till exempel i Vita och Svarta havet.
Bland de euryhalina arterna kom de vars ättlingar aktivt tog sig in i sötvattenskroppar. Processen att erövra floder och sjöar var gradvis. Först uppstod en grupp bräckvattenhydroider, som inte längre kunde återvända till havet, eftersom de inte kunde tolerera hög salthalt dess vatten Då kom bräckvattnen nära älvens mynningar. Inte alla av dem kunde övervinna denna "barriär", de flesta stannade vid flodens mynning. Cordylophora följer för närvarande denna väg.
Väl i floden mötte havsdjur en annan "barriär" på vägen - strömmen. När havs- eller bräckvatten-coelenterater aktivt trängde in i sötvatten, var de oundvikligen tvungna att övervinna det kommande vattenflödet, som förde planktonmaneter och fästa polyper eller deras kolonier tillbaka i havet. Förflyttningen av sådana fästpolyper mot flödet var svår.
I avlägsna geologiska epoker var kartan över jorden annorlunda än vi ser den nu. På många ställen var modern mark täckt av havet. När havet lämnade fanns slutna saltbassänger kvar, och havsdjur bevarades i dem. Några av dessa bassänger avsaltas gradvis och djuren antingen dog eller anpassade sig till de nya förhållandena. Det nu inneslutna Kaspiska havet, som i huvudsak är en enorm bräckt sjö, var en gång ansluten till havet och många djur av marint ursprung har bevarats i den. Bland dem finns en intressant coelenterat - Pallas merisia(Moerisia pallasi). Denna hydroidart har två former av polyper: vissa lever i en koloni på botten, andra leder en planktonisk livsstil. Flytande polyper bildar kolonier av två individer kopplade till varandra med sina ben. Då och då bryts kolonin på mitten, och på platsen för rasten utvecklar varje polyp en ny kronkrona, tentakel och mun. Dessutom förökar sig polyper också genom att de knoppas och separerar små fritt simmande maneter från sig själva. En närbesläktad art av Merizia lever i Svarta och Azovska havet, den andra i saltsjöarna i nordöstra Afrika.

Det är tydligt att alla tre arter av merisia härstammar från en gemensam förfader, som en gång levde i det gamla Sarmatiska havet. När Sarmatiska havet lämnade fanns ett antal vatten kvar på sin plats, inklusive det instängda Kaspiska havet och Egyptens sjöar. De utvecklade oberoende typer av Merizia.
Om du föreställer dig att avsaltningen av en reservoar går ännu längre, så kan du förstå hur sötvattensmaneter kan uppstå. Deras metod att erövra sötvattensbassänger är en långsiktig anpassning till ökad avsaltning. Samtidigt behöver de inte röra sig någonstans, de tar sig från havet till färskvatten inte i rymden utan i tiden.
1910 vid Atlantkusten Nordamerika Flera små vattenmaneter fångades. Det visade sig att de tillhörde en tidigare okänd art. Detta faktum i sig är inte särskilt betydelsefullt. Och nu beskrivs flera nya arter av coelenterater varje år - det finns fortfarande mycket outforskat i havet. En annan sak är intressant. Denna manet fick namnet blackfordia(Blackfordia) - 15 år senare fångades den i Svarta havet. Varken i Medelhavet, vars fauna är mycket välkänd, eller vid den europeiska kusten Atlanten denna art lever inte. Hur hamnade amerikanska blackfordia i Svarta havet? Den andra händelsen inträffade ganska nyligen. En av de typer av hydroider som lever i Kielkanalen är bougainvillea- upptäcktes oväntat igen i Svarta havet. Och blackfordia och nämnde Baltisk hydroid(Bougainvillia megas) - bräckvattenarter; för att ta sig från en bassäng med låg salthalt till en annan måste de, liksom Cordylophora, övervinna ett hinder - havet med dess höga salthalt.
Före byggandet av kanalen mellan Volga och Don fanns det bara två arter av coelenterates i Kaspiska havet - Kaspiska merisia och cordylophora. När kanalen var klar och navigeringen började flyttade ytterligare tre arter från Azov-Svartahavsbassängen till Kaspiska havet. Redan ett år efter att kanalen tagits i drift flyttade Blackfordia till Kaspiska havet, ett år senare Svarta havet Merisia och efter det den baltiska hydroiden (Bougainvillia megas), som kort innan hade gått in i Svarta havet från Kielbukten. Naturligtvis färdas inte bara coelenterater på detta sätt, utan också blötdjur, kräftdjur, maskar och andra bräckvattenorganismer.
"SEGELFLOTTA" AV CELINARITETER
Hydroid klassär uppdelad i två underklasser - hydroider Och sifonofor. Vi går vidare till beskrivningen av dessa fantastiska pelagiska koloniala coelenterater.
Hela världen av levande varelser lever på gränsen mellan två element - vatten och luft. På flytande alger, träfragment, pimpstensbitar och andra föremål kan du hitta en mängd olika fästa eller hårt klamrande djur. Man ska inte tro att de kom hit av en slump - de är "i nöd". Tvärtom är många av dem nära förbundna med både vatten- och luftmiljön och de kan inte existera under andra förhållanden. Förutom sådana "passiva passagerare" kan du här också se djur som aktivt simmar nära ytan, utrustade med olika utformade organ - flöten, eller djur som hålls på plats med en film av ytspänning av vatten. Hela detta komplex av organismer (pleiston) är särskilt rikt i subtroperna och tropikerna, där de destruktiva effekterna av låga temperaturer inte känns.
Ovan, när man diskuterade verkan av stickande celler, nämndes redan den "portugisiska krigsmannen" - en stor sifonofor physalia(Physalia, se färgskylt 8).

Liksom alla sifonoforer är physalia en koloni, som inkluderar både polypoida och medusoida individer. En luftbubbla, ELLER pneumatofor, stiger över vattenytan - en modifierad medusoid individ av kolonin. I stora exemplar når pneumatoforen 30 cm. Den har vanligtvis en ljusblå eller rödaktig färg. En luftbubbla flyter på havsytan som en hårt uppblåst gummiballong. Gasen som fyller den liknar till sin sammansättning luft, men har en högre halt av kväve och koldioxid och en minskad mängd syre. Denna gas produceras av speciella gaskörtlar inuti urinblåsan. Pneumatoforens väggar tål ganska starkt gastryck, eftersom de bildas av två lager av ektoderm, två lager av endoderm och två lager av mesoglea. Dessutom utsöndrar ektodermen ett tunt kitinoidskal, på grund av vilket styrkan hos pneumatoforen också ökar avsevärt, även om dess väggar förblir mycket tunna. Övre del Pneumatoforen har en åsliknande utväxt. Åsen ligger på pneumatoforen något diagonalt och har en lätt böjd S-form. Alla andra individer i kolonin är belägna på undersidan av pneumatoforen och är nedsänkta i vatten.
Matningspolyper, eller gastrozoider, sitter på en rad. De är mer eller mindre flaskformade och vända nedåt med munnen öppning. Varje matande polyp är utrustad med en lång tentakel - ett lasso. Hela längden av lassot är tätt täckt med stickande celler. Bredvid varje matande polyp, på undersidan av urinblåsan, är basen av gonodendron fäst - en individ av polypoidgenerationen. På gonodendra och dess laterala processer finns det kluster av reducerade medusoida individer - gonoforer, i vilka reproduktiva produkter utvecklas. Här sitter också skyddande tentakellösa polyper - palponer. Varje gonodendra har ett medusoidexemplar som kallas en nektofor eller simklocka. Reproduktionsceller bildas inte i nektoforen, och dess paraply når en betydande storlek och kan dra ihop sig, som hos fritt simmande maneter. Innan gonoforernas sexuella mognad börjar, lossnar gonodendra från kolonin och simmar vid havets yta, med nektoforen som utför rörelsefunktioner.

På grund av det sneda arrangemanget av åsen på simblåsan är physalia asymmetrisk, och två former av physalia är kända - "höger" och "vänster", som så att säga är en spegelbild av varandra. Det märktes att alla physialia som bor i ett område av havet har samma struktur, det vill säga att de alla är antingen "höger" eller "vänster". I detta avseende har det föreslagits att det finns två arter eller två geografiska raser av physalia.
Men när de började studera utvecklingen av dessa sifonoforer upptäcktes det att bland avkomman till en fysalia finns det alltid lika många "höger" och "vänster". Det betyder att Physalia inte har några speciella raser. Men hur uppstår kluster av "vänster" och "höger" sifonoforer och varför förekommer inte dessa två former tillsammans?
Svaret på denna fråga erhölls efter en detaljerad studie av strukturen av luftblåsan av physalia. Det visade sig att formen och placeringen av åsen i dess spets är mycket viktiga för physalia. Som nämnts ovan är physalia-ryggen något böjd i form av bokstaven S. Physaliaen rör sig längs havsytan på grund av att vinden träffar dess luftbubbla. Om det inte fanns någon ås skulle sifonoforen ständigt röra sig i en rak linje och så småningom spolas iland. Men närvaron av en ås gör betydande förändringar i seglingsriggen för "den portugisiska krigsmannen". En snett ställd och krökt krön tvingar djuret att simma i spetsig vinkel mot vinden och då och då göra en sväng runt sin axel mot vinden.
Om du observerar en physalia som simmar nära stranden, i vars riktning vinden blåser, kan du se hur den antingen närmar sig stranden, och sedan oväntat vänder den andra sidan till observatören, simmar den långsamt bort från honom. Hela armadas av "portugisiska fartyg" manövrerar på detta sätt, som påminner om segelflottans agerande under de medeltida krigen. När de rör sig beter sig de "höger" och "vänstra" "portugisiska båtarna" olika. Under påverkan av vinden som blåser i en riktning divergerar de i olika riktningar - "höger" till vänster och "vänster" till höger. Det är därför kluster av identiska former av fysalia uppstår.
Pleistoniska organismer inkluderar också mycket märkliga coelenterater - porpita(Porpita) och velella(Velella), även kallad segelfisk.
Under lång tid klassificerades dessa djur som sifonoforer, och deras individuella bihang ansågs vara specialiserade individer av kolonin. Nu är allt fler zoologer benägna att tro att porpita och swallowtail inte är en koloni, utan en stor enkel flytande polyp, och klassificerar dem som ordning kondrofora(Chondrophora) från hydroid klass. Deras kropp är tillplattad; hos porpita har den formen av en cirkel, hos segelfisken har den formen av en oval. Den övre sidan av skivan är täckt med ett kitinoidskal, under vilket en komplex luftklocka placeras - en pneumatofor. Den består av en central kammare, ett stort antal ringkammare som omger den och tunna rör som sträcker sig från dem till alla delar av kroppen - luftstrupen, som tjänar till andning. Polypens organ är belägna på undersidan av skivan. I mitten finns en munkon, och längs periferin finns det många tentakler. Mellan munkonen och tentaklarna finns speciella utväxter av kroppen - gonodendra, på vilka individer av medusoidgenerationen knoppar. Den övre sidan av skivan av kustnära porpita är slät; Velella, som lever i det öppna havet, har en hög triangulär utväxt på sig - ett segel. Velellas seglet har samma betydelse som krönet på physalias luftblåsa. Den är placerad på segelbåtens ovala kropp asymmetriskt och lätt S-formad. Seglet tillåter djuret att inte röra sig i en rak linje, utan att manövrera, även om det naturligtvis inte är godtyckligt, utan mer eller mindre slumpmässigt.
I subtropiska delar av havet, där temperaturen inte sjunker under 15°C, finns segelfiskar i mycket stora mängder. På vissa ställen samlas dessa stora coelenterates (de når 12 cm längs skivans långa axel) i enorma flockar som sträcker sig flera tiotals mil, och för varje kvadratmeter havets yta faller på segelbåten. Unga segelfiskar, vars storlek mäts i millimeter, simmar också tillsammans med stora segelbåtar.
Vinden, som slår mot seglet, driver en flock velella över havet, och de kan resa många hundra mil.
När de bor i det öppna havet är segelbåtar inte rädda för vatten: de kan inte drunkna, eftersom de har en mycket avancerad pneumatofor, som består av ett stort antal oberoende kammare. Om en våg ändå välter välellan, återgår den, med hjälp av rörelser av skivans kanter, till sitt normala läge och utsätter seglet igen för vinden. Här kan man förutom segelbåtar även hitta många andra djur, som dock är nästan osynliga till en början.
Det är välkänt att det öppna havet i tropikerna har en intensiv blå färg. I detta avseende är segelbåtar och de flesta av djuren som lever med dem också färgade blå eller blå - detta fungerar som ett bra skydd för dem.
Segelbåtar och andra djur som lever bland dem skapar en speciell, nära sammankopplad värld i det öppna havet - en pleistonisk biocenos, som av strömmens och vindens vilja ständigt flyter på havets yta.
Velella, som alla coelenterater, är ett rovdjur; den livnär sig på plankton, dess föda inkluderar kräftdjur, larver av olika ryggradslösa djur och fiskyngel. Alla andra djur som ingår i den flytande biocenosen livnär sig antingen på segelbåtar eller använder dem som ett permanent eller tillfälligt substrat för fastsättning. Alltså existerar hela biocenosen på bekostnad av plankton, men bara segelfiskar använder direkt plankton.
Små blå krabbor färdas på översidan av velellaskivan, som på däcket på ett fartyg. planer(Flygplan). Här hittar de skydd från fiender och får även mat. En hungrig krabba rör sig snabbt till undersidan av segelfiskens skiva och tar bort de fångade planktoniska kräftdjuren. Efter att ha ätit klättrar krabban igen upp på skivans ovansida och slår sig ner under seglet och klänger sig tätt intill det. Krabbor slukar aldrig sitt skepp, vilket inte är fallet med många andra pleistoniska djur.
På undersidan av segelfisken kan man ofta hitta rovsnäckan Janthina. Yantiner äter mjuk vävnad tills bara ett kitinoida skelett finns kvar från segelbåten. Efter att ha förlorat stödet sjunker yantina inte, eftersom den är väl anpassad till livet på vattenytan. Så snart svalstjärten som äts börjar sjunka, släpper yantinen rikligt med slem och bildar bubblor fyllda med luft. Detta slem hårdnar mycket snabbt, och ett bra flyt erhålls, på vilket blötdjuren självständigt kan simma och flytta från en segelbåt till en annan. Simma till nytt offer, Yantina lämnar flottören som nu är onödig för henne och kryper snabbt upp på velellan. Den övergivna yantinaflottan befolkas snart av hydroider, mossor, havstulpaner och andra vidhäftade djur, såväl som små krabbor; ibland sätter de sig på själva blötdjurets skal.
Tillsammans med jantinope bosätter sig också ett annat rovdjur, nakensnäckan Aeolis, på segelbåtar.
Ibland kan man, bredvid segelfisken, se de medföljande nakensnäcka blötdjuren (Glaucus). Kroppen på denna skallösa blötdjur är långsträckt, fiskformad, på sidorna finns tre par grenade tentakelliknande utväxter, med vars hjälp blötdjuret fäster på vattenytans film. Den simmar med sin mörkblå buksida uppåt, ryggsidan är silvervit. Detta gör att simglaucusen blir osynlig både från luften och från vattnet. En hungrig glaukus, som krattar upp med tentakelliknande utväxter, simmar fram till segelbåten och håller i den, drar sig ut och äter upp stora bitar av skivans kant.
När de äts av blötdjur dör segelbåtarna, men kvar är ett kitinoida skelett, i vilket systemet av luftkammare fortfarande finns bevarat. Sådana döda segelbåtar flyter på ytan under en tid, och larverna av havstulpaner (Lepas fasciculatus) sätter sig på dem. När de nya bosättarna växer sjunker segelfiskens skelett djupare och djupare, och på benet, med vars hjälp ankan fästs på underlaget, utvecklas ytterligare en sfärisk flöte, vilket ökar kräftdjurets flytförmåga.
Alla frilevande havstulpaner är fästa djur, med det enda undantaget som ovan nämnda arter av havstulpaner. När dess sfäriska flottör når en betydande storlek, separeras den från segelbåten, och efter det kan sjöankan självständigt flyta på vattenytan och till och med simma och svänga med benen. I andra havstulpaner driver flaxandet av benen maten mot kräftdjuret - små planktoniska organismer, men denna art av havstulpan, till skillnad från alla sina släktingar, leder en rovdjursstil. När den simmar fram till segelbåten, tar sjöankan tag i kanten på sin skiva med sina ben och äter snabbt bort en betydande del av velellan när den rör sig längs kanten.
Utöver de djur som beskrivs här omfattar velellabiocenosen även en del räkor, ögonfransmaskar, vattenstrider och ett antal andra djur, inklusive en art av flygfisk, Prognichthys agae, som lägger ägg på segelbåtar. Halobater vatten strider buggar lever i nära kontakt med Velella och Porpita och använder dem både som en "paj" och som en "flotte".
Velellas värld som flyter i det öppna havet är mycket begränsad, men alla dess invånare är nära förbundna med varandra. Det är intressant att notera att de flesta av de arter som utgör denna biocenos tillhör grupper av djur som vanligtvis leder en bottenlevande livsstil. Utifrån detta kan vi med tillförsikt säga att pleistoniska djur kommer från bentiska (och inte planktoniska) organismer som tappade kontakten med botten och började fästa vid olika flytande föremål eller använda vattenytans film som stöd.
Djurliv: i 6 volymer. - M.: Upplysning. Redigerad av professorerna N.A. Gladkov, A.V. Mikheev. 1970 .
- - (Hydrozoa) klass av vattenlevande ryggradslösa djur såsom coelenterata (Coelenterata). De flesta G. kännetecknas av generationsväxling: Polyper ersätts av den sexuella generationen av maneter (Se maneter). De flesta G. har en asexuell generation... ... Stora sovjetiska encyklopedien
ALLMÄNNA EGENSKAPER Coelenterates är de mest dåligt organiserade av de sanna flercelliga djuren. Kroppen av coelenterates består av två lager av celler, ektoderm och endoderm, mellan vilka det finns mer eller mindre... ... Biologisk uppslagsverk
I moderna system klassificeringar, är djurriket (Animalia) uppdelat i två underriken: parazoer (Parazoa) och äkta flercelliga organismer (Eumetazoa eller Metazoa). Endast en typ av svamp klassificeras som parazoan. De har inte riktiga vävnader och organ... ... Colliers uppslagsverk
Turritopsis ... Wikipedia
Hydroidolina ... Wikipedia
Obelia sp ... Wikipedia
Bathykorus bouilloni (Aeginidae) ... Wikipedia
Den här artikeln handlar om havsdjur. För att kasta vapen, se Siphonophore. Sifonoforer ... Wikipedia
- I kontakt med 0
- Google+ 0
- OK 0
- Facebook 0